医学免疫学知识总结
第三章 抗原(Antigen,Ag)
一 . 基本概念
1.抗原(Antigen,Ag):是一类能刺激机体免疫系统使之产生特异性免疫应答,诱导产生抗体及效应T淋巴细胞;并能与之在体内体外发生特异性结合的物质。又称免疫原。
2 .免疫原性:刺激机体免疫系统,使之增殖、分化,产生抗体及效应淋巴细胞的特性。
3 .抗原性:能与抗体及效应淋巴细胞发生特异性结合的特性。
4 .完全抗原:既有免疫原性又有抗原性的物质。
5. 半抗原:只有抗原性没有免疫原性的物质。
二. 决定免疫原性的因素
(一)抗原方面的因素
1 .异物性:是免疫原的核心。抗原与机体亲缘关系越远,组织结构差异越大,免疫原性越强。
免疫系统识别自身与非己的本质:胚胎期或未成熟免疫细胞发育期遇到的所有抗原,包括胚系及非胚系基因编码的产物为自身物质。免疫细胞未成熟期未遇到的物质为非己物质。
2 . 抗原分子的理化特性:
(1)化学组成及异质性:要求一定的化学复杂性。蛋白质是良好的抗原。
(2)分子量:10kD以上具有抗原性,100kD以上为强抗原。分子量大,抗原决定基越多。
(3)结构的复杂性:直链氨基酸组成的蛋白质稳定性差,加入苯环氨基酸后,免疫原性加大。分子构象、物理状态等对免疫原性都有影响。
3. 可递呈性:诱导细胞免疫反应所要求。
(二)生物学方面的因素
1 .宿主的遗传背景:相同的抗原在不同动物所诱导的免疫应答有明显的差异,这与动物的MHC背景有关。
2 .年龄、性别与健康状态的影响。
3. 引入抗原的剂量、途径与次数:剂量应适中;途径依次为皮内--皮下--腹腔、
静脉--口服;间隔适当。
三. 抗原表位(抗原决定簇)
抗原决定簇(Antigen Determinant):
抗原分子中决定抗原特异性的特殊化学基团。是被免疫细胞识别的靶结构或基本单位,又称表位(Epitope)也是特异性免疫反应的物质基础。
表位的性质、数目、空间结构决定了抗原的特异性。T、B细胞识别抗原上的不同抗原表位,故有T细胞表位及B细胞表位之称。
1 . B细胞表位的特性:
能识别天然抗原分子的表位,一般为不连续的、经三维折叠后空间构象上聚集形成的基团(构象表位),或抗原分子表面连续的氨基酸片段(长链弯曲折叠处,顺序表位)。
2 .T细胞表位的特性:
只能识别经APC处理过的,经MHC传递的小分子多肽(镶嵌在MHC分子的凹槽中)。它由序列上相连的氨基酸(顺序表位)组成,在天然分子中是位于抗原分子内部的疏水基团。
四. 抗原的种类
1. 根据抗原是否显示免疫原性区分:完全抗原、半抗原
2. 根据B细胞产生抗体是否需要T细胞参与而区分: 胸腺依赖抗原(TD-Ag) 非胸腺依赖抗原(TI-Ag)
3. 根据抗原与机体的亲缘关系而区分:异种抗原、同种异体抗原、自身抗原、异嗜性抗原
五. 淋巴细胞激活剂
指一类能使高比例活化淋巴细胞的物质,但它们不属于抗原,因为它们对细胞克隆的刺激不涉及抗原特异性。
1. 丝裂原:
包括多种成分。如植物凝集素(ConA、PHA、PWM),是一类含蔗糖的蛋白质,可以和各种细胞表面的糖蛋白结合,通过信号传递引起细胞活化、增殖。 常见的淋巴细胞激活剂
种类 英文缩写 来源 类别 靶细胞
刀豆素A ConA Jack豆 凝集素 T细胞
植物血凝素 PHA 菜豆 凝集素 T细胞
美洲商陆 PWM 美洲商陆 凝集素 T细胞
脂多糖 LPS G-菌 内毒素 B细胞
2. 佐剂:
与抗原一起用于机体,可增强免疫应答或改变应答类型,但不影响特异性。其机制有 :1)增强抗原的滞留时间。2)增强APC细胞对抗原处理、提呈能力。3)刺激淋巴细胞增殖、分化等。
包括卡介苗(BCG)、脂多糖(LPS)、细胞因子(如GM-CSF)、聚肌胞(Poly I:
C)、寡核苷酸CpG、福氏佐剂(CFA、IFA)等。
3. 超抗原(SAg):
某些抗原物质,只需很低的浓度即可激活2%-20%T细胞克隆活化,产生极强的免疫应答,但有不同于丝裂原的作用,称为超抗原(Superantigen,SAg )。 超抗原有两类:外源性超抗原---如一些细菌的外毒素;内源性超抗原---如一些病毒感染后表达在细胞上的病毒蛋白。
还有活化T细胞超抗原及活化B细胞超抗原之分。
超抗原活化T细胞的机制:与T细胞表位一样,形成MHC-SAg - TCR结构,所不同的是,它无需APC的处理,只是附着在MHC凹槽的外侧;与TCR的结合也不涉及特异性,而是和TCR链的V区结合
第五章 补 体 系 统 (complement system)
一 . 基本概念
1、 补体:是存在人和动物血清中的一组(30余种)具有酶活性的、不耐热的蛋白质,它可协助抗体完成对病原体的裂解清除。
2、补体的激活:自然条件下,补体在血浆中以无活性的酶原形式存在,多种特异性及非特异性免疫学机制可以使之分解,产生有活性的分子,补体成分的激活是一个连锁反应,最终结局导致其所附着的细胞的溶解。
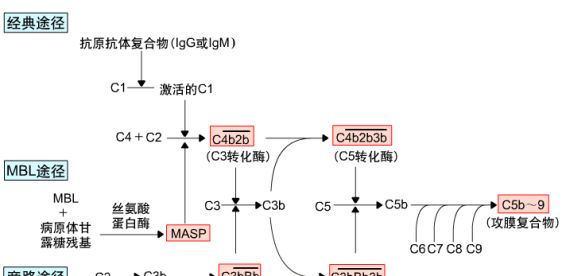
二. 补体的激活途径:
包括经典途径、BML(甘露聚糖结合凝集素)途径及C3旁路途径。
三条途径的起始阶段不同,但具有共同的末端通路---攻膜复合体的形成及细胞溶解效应。
1. 经典激活途径
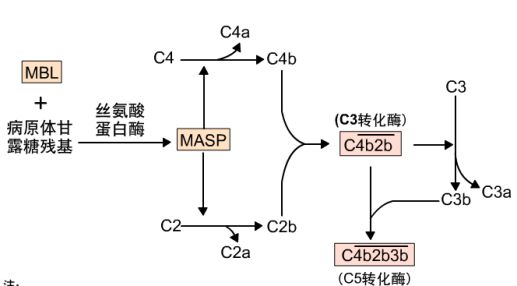
2. MBL激活途径
发生在微生物感染的早期,一些细胞因子使肝细胞产生急性期蛋白,其中包括MBL,可与甘露糖残基结合。细菌细胞表面有甘露糖残基暴露,而脊椎动物细胞表面则是被其他糖基覆盖。

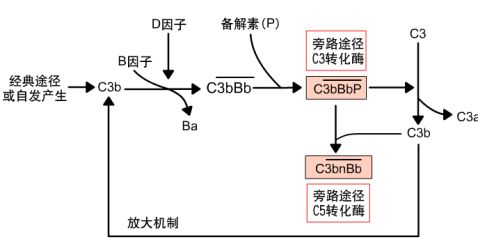
3 .旁路激活途径
4. 三条激活途径的共同末端效应
三. 补体活化的调控
1. 补体的自身调控:
激活过程中生成的某些中间产物极不稳定,成为级联反应的重要自限因素。如C4b2b、C3bBb等。
2.调节因子的作用:有十余种,按作用可以分为三种:
1)防止或限制补体在液相中自我激活。
2)抑制或增强补体对底物正常作用。
3)保护机体组织细胞免遭补体破坏。
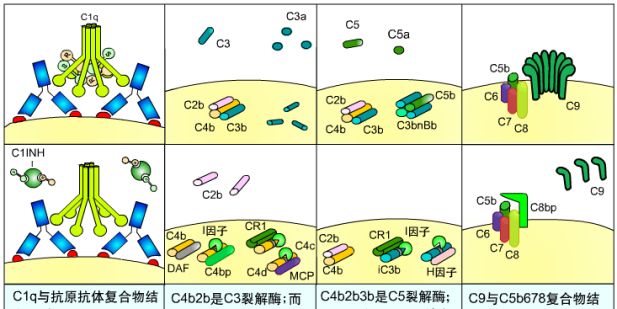
几种补体调节因子的作用:
1)C1抑制物(C1INH):与C1r、C1s结合,使它们失去酶解正常底物的能力。
2)C3转化酶抑制剂:C4结合蛋白与CR1--与C4b结合;I因子--裂解C4b;膜辅蛋白(MCP)--促进I因子的作用;衰变加速因子(DAF)--同C2竞争地与C4b结合;H因子--同Bb竞争性地与C3b结合
3)保护细胞膜:C8结合蛋白--干扰C8、C9的结合;膜反应性溶解抑制物(MIRL)--阻止C7、C8与C5b6的结合。 S蛋白--阻止C5b67插入胞膜。
几种补体调节因子作用模式图
四 补体受体
1. 补体裂解片段受体:
1)补体受体1(CR1,C3b/C4bR)--表达在许多细胞表面。有抑制补体激活、调理、清除免疫复合物、免疫调节的作用。
2)补体受体2(CR2,C3dR,CD21):存在于B、DC表面,有免疫调节作用。 3)补体受体3(CR3)、补体受体4(CR4)
2. 可溶性C3a/C4a/C5a受体:
表达在肥大细胞、嗜碱性粒细胞等细胞表面, C3a/C4a/C5a与其结合后可介导炎症反应,以C5a作用最强。
3. 调节补体级联反应的受体:如H因子\MCP\DAF等分子的受体。
五、补体的生物学活性
1. 补体介导的细胞溶解(CDC) 2. 调理作用
3. 参与炎症反应 4. 清除免疫复合物
5. 免疫调节作用
6. 与其他酶系统相互作用
第六章 细胞因子(Cytokine,CK)
第一节 细胞因子的概述
一、细胞因子的概念
是各种细胞分泌的具有生物活性的小分子蛋白物质的统称。主要作用为介导各类免疫细胞间的相互作用。在非特异性免疫及特异性免疫应答,细胞生长分化、调节免疫功能、炎症发生和创伤愈合中发挥重要功能。
可以产生细胞因子的细胞有:
免疫细胞---单核巨噬细胞,T、B淋巴细胞,NK细胞
非免疫细胞---血管内皮细胞、表皮细胞、成纤维细胞等。
二、细胞因子的共同特性
1、低分子量(15--30kD,多为25kD以下)的糖蛋白。多以单体存在。
2、以配体--受体的方式发挥作用,有较强的亲和力,微量的细胞因子即可产生很强的生物学效应(pM)。作用的方式为非特异性,即对靶细胞作用无抗原特异性,也不受MHC限制。
3、在抗原、丝裂原和其他活性物作用下而分泌,过程短暂、呈自限性。半衰期也很短。
4、作用的方式有三种--- 自分泌:靶细胞与分泌细胞为同一细胞。 旁分泌:靶细胞与分泌细胞非同一细胞,但
二者邻近。 内分泌:靶细胞与分泌细胞非同一细胞,二者相距较远。
5、作用的特点--- 多效性:一种因子作用在不同的靶细胞可以产生不同的生物学作用。 重叠性:不同的细胞因子可对同一靶细胞发挥相同的生物学效应。 拮抗效应:两种细胞因子相互作用时,其中一种削弱另一种的作用。 协同效应:两种细胞因子同时存在时,彼此的作用得到加强。 双向性:适量时具有生理调节作用,过量则产生对机体的损伤。
6、细胞因子的网络性:无论是产生、发挥作用、受体表达、相互调节等均存在网络特点。
三、细胞因子的分类(按结构及受体结构)
1、造血因子家族(Hematopoietin):
参与造血细胞的生长及分化。结构上、氨基酸组成差异很大,但立体结构上相似---都含有4个螺旋的基本结构。
成员:多种白介素(IL-2、3、4、5、6、7、9、11、12、15)
集落刺激因子(GM-CSF、G-CSF、M-CSF 、EPO )
2、干扰素家族(Interferon,IFN):
干扰病毒感染及复制的作用。有 IFNα、 IFNβ、 IFNγ。
I型干扰素: IFNα、β,由白细胞、成纤维细胞及病毒感染的组织细胞产生。 II型干扰素:IFNγ,由活化的T细胞和NK细胞产生,也称免疫干扰素。
3、肿瘤坏死因子家族(Tumor necrosis factor,TNF):
分TNF α、TNFβ两类。前者由单核巨噬细胞、活化的T细胞、NK细胞产生;后者由活化的T细胞产生,又称淋巴毒素。
4、趋化性细胞因子家族(Chemokine):
由十余种在结构上有较大同源性、分子量多为8-10kD的蛋白组成。根据在氨基端半胱氨酸的排列及含量,又可区分为四个亚家族:
α亚家族---CXC,如IL-8,趋化中性粒细胞;
β亚家族---CC,如MCP-1,趋化单核细胞;eotaxin趋化嗜酸性粒细胞。 γ亚家族---C,如淋巴细胞趋化蛋白,对淋巴细胞有趋化作用。
δ亚家族---CX3C,如神经趋化素,介导单核细胞/淋巴细胞与产生它的细胞间的粘附。
5、生长因子家族(Growth Factor,GF):
具有刺激细胞生长作用的因子的总称。
包括转化生长因子β(TGFβ) 、表皮细胞生长因子(EGF) 、血管内皮生长因子(VEGF) 、成纤维生长因子(FGF) 、神经生长因子(NGF) 、血小板衍生生长因子(PDGF)等。
第二节 细胞因子的生物学活性
一、介导和调节天然免疫
单核巨噬细胞分泌的细胞因子。作用---
1、抗病毒的细胞因子:I型干扰素、IL-12、IL-15。
2、前炎症细胞因子:TNF、IL-1、IL-6和趋化性细胞因子。
活跃在天然免疫中的几种细胞因子---
A、刺激细胞合成多种酶,干扰病毒RNA或DNA的复制。
B、增强NK细胞对病毒感染细胞的裂解作用。
C、刺激被病毒感染的细胞的MHC--I类分子表达,增强CTL的活性。
靶细胞是T细胞、NK细胞,是连接天然免疫与特异性免疫的纽带,主要提高细胞免疫功能。
A、刺激T细胞、NK细胞分泌IFN-γ。
B、促CD4向Th1分化,提高巨噬细胞的功能。
C、增强CTL细胞和NK细胞的杀伤功能。
单核巨噬细胞、T、NK细胞、肥大细胞产生, IFNγ增加其合成。
A、刺激血管内皮细胞表达粘附分子,促白细胞粘附于内皮细胞表面。
B、刺激单核巨噬细胞分泌趋化因子,招募白细胞至炎症部位。
C、激活炎性细胞杀灭病原体。
D、参与组织修复,诱导血管和结缔组织形成。
E、刺激单核巨噬细胞、血管内皮细胞产生IL-6、IL-1。
IL-8):中性粒细胞;
MCP-1):单核细胞; Eotaxin趋化嗜酸性粒细胞; 还有T细胞、NK、DC、嗜碱性粒细胞趋化因子。
:淋巴细胞
:在脾及心脏表达,介导单核细胞/淋巴细胞与产生它的细胞间的粘附。
趋化因子的最基本作用:趋化白细胞的游走至炎症部位。
实质是吸引白细胞向浓度逐渐增高的趋化因子部位移动;趋化因子浓度梯度的形成与基质表面、内皮细胞表面的粘蛋白浓度有关。
1)IL-1:刺激单核巨噬细胞、内皮细胞分泌趋化因子,刺激血管内皮细胞表达粘附分子。
2)IL-6:刺激肝细胞分泌急性期蛋白。
3)IL-10:是天然免疫中重要的负调节因子.可以抑制巨噬细胞分泌IL-1、IL-6。
二、介导和调节特异性免疫应答
由抗原激活的T淋巴细胞产生分泌,作用在淋巴细胞的激活、生长、分化及效
应各个阶段。
1、刺激APC表达MHC分子:IFN
2、促CD4活化:IFN
3、促免疫活性细胞分化增殖:L-12促CD4Th0--Th1,IL-4促Th0--Th2
4、效应阶段,促免疫细胞对抗原物质进行清除:
A、Th1 产生的IFN-γ、 IL-2,激活单核巨噬细胞、NK、CTL细胞---细胞免疫;
B、Th2产生IL-4、IL-5,刺激B细胞、嗜酸性粒细胞分化,促抗体产生,促杀伤蠕虫,介导变态反应。
5、B细胞在产生抗体过程中,发生抗体类别的转换,也是在细胞因子的作用下实现的。细胞因子介导的B细胞分化、增殖及抗体类别转换
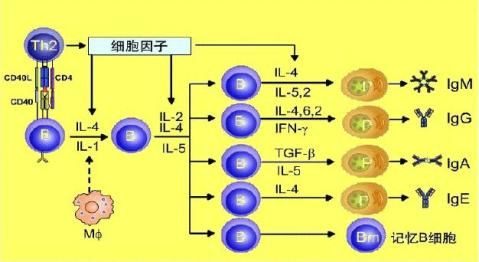
参与特异性免疫应答的细胞因子
主要由CD4(Th1)、CD8 T细胞产生,以自分泌及旁分泌方式产生效应,其作用沿种系向上有 约束性,向下无约束性。是T细胞激活的关键因子(TCGF)。另外对B细胞、NK细胞、单核巨噬细胞都有激活及促增生的作用。
CD4(Th2)细胞 产生,是B细胞的生长分化因子:促B细胞MHC-II表达、促抗原提呈、促静止B细胞表达FcR、促活化B细胞产生IgE/IgG1。与体液免疫、I型变态反应有关。
T细胞和NK细胞产生。生物学活性有明显的种属特异性,除有抗病毒活性外,还有独特的功能---
A、是单核巨噬细胞的主要激活、活化因子。
B、诱导多种细胞 表达MHC I、II类分子。
C、促Th0--Th1转化,促CTL细胞成熟。
D、促抗体类别向IgG2a、IgG3转换,它们有强的补体激活及免疫调理作用。同时还可抑制向IgG1、IgE的转换。
E、激活 NK、中性粒细胞、血管内皮 细胞。
与免疫功能相关性强的是TGF- β1,它主要由T淋巴细胞(Th3)及单核细胞产生。在功能上拮抗淋巴细胞反应,抑制淋巴细胞增殖,抑制CTL、巨噬细胞的作用,因此认为它是一种免疫系统关闭的信号 。其产生细胞被认为是抑制性细胞。肿瘤细胞可以通过产生大量的TGF- β来逃避免疫系统的作用。
三、刺激造血细胞生成和分化
第三节 细胞因子受体
根据受体的结构,可将其分为五个家族:
I型细胞因子受体家族(造血因子受体家族)
II细胞因子受体家族(IFN受体家族)
III细胞因子受体家族(TNF受体家族)
Ig基因超家族
趋化因子受体家族(七次跨膜受体)
一、细胞因子受体的共同特征:
1、都是跨膜蛋白,由膜外区、跨膜区、胞浆区三部分组成。
2、膜外区由一个或多个结构域组成。结构域有三种类型:细胞因子结构域(CK)、 III型纤连蛋白型结构域(FNIII)、Ig样结构域
3、多亚单位受体及受体共用亚单位:从功能角度可将受体分为结合亚单位与传递亚单位两部分。不同的CKR的结合亚单位结构各异,传递亚单位结构变化小,这是不同的细胞因子具有相同生物学活性的基础。将相同的传递亚单位称共用亚单位。
二、CKR介导的信号传导:
1、酪氨酸激酶(PTK)介导的信号传导途径:有的受体胞内区带有PTK结构,而有的受体胞内区未带有PTK结构域,但可结合有PTK家族的成员。与受体结合的PTK多见Janus家族成员(Jak成员),一个亚单位可连接一个或多个Jak家族成员。CK--CKR结合致使JaK成员相互靠拢,磷酸化激活,启动多条信号传导通路。其中Jak-State通路最重要。
Jak Y)---磷酸化(pY) StatSH2结构域---磷酸化
Stat
2、G蛋白偶连受体介导的信号传导:
G蛋白为三聚体结构( α 、β、γ), α亚基上有GDP、GTP位点。
CK结合 暴露出α亚基上的结合位点 进行GDP-GTP的转换 α亚基与β、γ 暴露出α 被激活并 产生大量的cAMP同时α亚基恢复原有构象 与腺苷酸环化酶解离 、γ亚基再结合。
第五节 细胞因子及相关制剂在临床的应用
一、感染性疾病:
1、细菌性脓毒血症休克(BSS):
LPS刺激巨噬细胞产生过量的IL-1、TNF引起。给予IL-1受体拮抗剂和TNF单克隆抗体可降低死亡率。
2、病毒性感染:
干扰素、IL-12已用于HIV感染,纠正病人Th1细胞的进行性减少。
二、肿瘤:
IL-2活化NK、T细胞--LAK细胞、TIL细胞;IL-2、TNF、IFN等与瘤苗一起应用,增强其诱发的免疫反应;CSF用于放疗后的粒细胞减少。
三、器官移植:
用细胞因子拮抗剂或可溶性受体抑制对移植物的排斥,延长器官的存活时间,如IL-1R、IL-2R、交连毒素的CK等。
四、过敏反应:
IL-4、IL13拮抗剂可减少IgE的产生 。IFN-有相同的作用。
五、自身免疫性疾病:
如IL-10治疗由Th1引起的自身免疫性疾病。TNF中和抗体减轻类风湿性关节损伤。
第六节 细胞因子极其受体的检测
一、细胞因子检测的方法
1、免疫学检测法:
其基本原理是将细胞因子作为抗原进行定量检测。如免疫斑点法、ELISA法、RIA法和免疫印迹法等均已用于细胞因子的检测。
2、生物学测定法:
其原理是根据细胞因子对特定的依赖性细胞株(即靶细胞)的促增殖作用,以增殖细胞中的DNA的合成或酶活性为指标,间接推算出细胞因子的活性单位。
3、分子生物学测定法:目前采用的有RNA印迹法、原位杂交、PCR等,还可通过检测mRNA表达量,推算CK的合成量。
二、CKR检测的基本技术
1、活细胞吸收实验 :将过量的待检细胞与限量细胞因子(配基)共孵育,若细胞膜表面存在相应的受体即可吸收配基,通过测定回收后配基生物活性的丧失情况可确定受体是否存在。
2、同位素标记重组配基的放射受体分析:这是确定细胞受体分布及其特性的主要方法,可测定受体的数量和亲和力。一般采用重组细胞因子体外标记同位素。
3、抗受体McAb:利用McAb即可通过封闭受体抑制相应细胞因子的生物活性,也可用标记的McAb直接作免疫放射受体分析或免疫沉淀。
4、重组细胞因子与受体的交联分析:利用化学交连剂将标记的重组CK与膜受体交连,细胞裂解物经PAGE后做放射自显影,通过带型分析确定受体的分子量几亚单位。
5、受体cDNA分析:通过核苷酸序列推导的氨基酸序列并进行结构功能区分析。
6、可溶性CKR的检测:某些CKR除了存在于细胞膜外,还可分泌进入体液,用sCKR表示,可以用免疫学的方法来检测。
第七章 白细胞分化抗原
第一节 白细胞分化抗原及有关概念
1、细胞表面标记:
存在于免疫细胞膜表面、参与免疫细胞间相互识别的膜分子的统称。如MHC、CD、TCR、BCR、FcR、CKR和其他分子(粘附分子)。
2、白细胞分化抗原:
指白细胞在分化、成熟为不同谱系、分化的不同阶段,以及细胞活化过程中出现或消失的细胞表面标记分子。成分是蛋白质或者糖蛋白。
3、CD(cluster of differentiation,分化群):
应用以单克隆抗体鉴定为主的聚类分析方法,将来自不同实验室的单克隆抗体所识别的同一分化抗原归为一个分化群(CD)。CD即位于细胞膜上一类分化抗原的总称。
4、类别:
1)已确定的CD 分子共有166种(19xx年的资料)。大致可分为9个组:T细胞、B细胞、髓系细胞 、NK细胞、血小板、内皮细胞、粘附分子、细胞因子
受体、非谱系 。
2)根据白细胞分化抗原胞外区结构特点,分为不同家族,包括---免疫球蛋白超家族、细胞因子受体家族、TNF超家族、TNF受体超家族、C型凝集素超家族、整合素超家族等。
第二节 参与T细胞抗原识别与活化的CD分子
T细胞是重要的免疫活性细胞,在免疫反应中处于中心地位。与它接触的细胞有APC-T、T-B、T-靶细胞。其中,重要的CD分子有:、、等。
1、CD3:
CD3分子与TCR共同组成TCR/CD3复合体,分布在T细胞的标志。CD3可将TCR识别的信号传入细胞内,引起T细胞的活化。 TCR是由两条链组成的二聚体,有TCRαβ、TCR γδ两种形式。跨膜区为带正电荷的氨基酸组成。
CD3分子由γ、δ、ε、δ、ε 5条链组成,呈γ ε 、δ ε、 δ δ 、 δ ε二聚体形式,跨膜区氨基酸带负电荷,与TCR所带的正电荷通过盐桥形成稳定的复合体。
CD3分子的胞浆区内有“免疫受体酪氨酸活化基序”(ITAM)结构,其中的酪氨酸磷酸化后,可通过活化有关激酶,传导引起T细胞活化的信号。 2、CD2(SRBC、LFA-2)
2SRBC:绵羊红细胞受体
LFA-2 :淋巴细胞功能相关抗原-2
CD2为B细胞无),还表达在胸腺细胞、NK细胞。其分子为一条由327个氨基酸组成的跨膜肽链,胞外区结构类似与Ig的V区、C2区,胞浆区富含脯氨酸,是与多种蛋白酪氨酸激酶(PTK)相连的区域。
CD2的配体是CD58(LFA-3)。
3、CD58 (LFA-3)
CD58属Ig超家族,结构与CD2相似。存在人、绵羊的红细胞膜表面,是玫瑰花环形成试验的结构基础。CD58与CD2结合的生物学作用:
1)促进T细胞与APC粘附、增强TCR识别,参与信号传递的。
2)与胸腺细胞分化成熟有关,胸腺发生MHC-Ag-TCR结合的T细胞,克隆被
淘汰。而MHC-Ag-TCR不能结合的T细胞,通过CD2-CD58结合促其成熟。 4、CD4:
为单链跨膜糖蛋白,胞膜外区属IgSF家族,有四个结构域;胞质区有与PTK结合的基序。有CD4的T细胞为T辅助(Th)细胞。CD4分子通过胞外第一、二结构域与MHC II类分子的非多肽区结合,辅助TCR-CD3对抗原的识别;通过胞质区与PTK P56的结合,参与信号转导。
5、CD8:
由两条链构成, α、β链胞外区各有一个V样结构域(IgSF), 有αα同聚体、αβ异聚体, 胞质区与PTK相连。
CD8分布在成熟(CTL)及未成熟T细胞(CD4、8双阳性)上,是TCR识别MHC -I类抗原的辅助受体,并参与CTL细胞活化信号的传递。CD8结合的部位为MHC-I的α3结构域。
6、CD28/CTLA-4(CD152):
均为两条多肽链组成的同源二聚体,有31%的序列同源,配基都为B7-1(CD80)/B7-2(CD86)。
CD28存在于几乎所有的CD4细胞及50%的CD8细胞上,部分浆细胞及B细胞也可表达,作为辅助刺激分子,提供T细胞活化的第二信号。是T-B细胞协作的重要的分子基础。
CTLA-4表达在活化的T细胞膜上,通过胞浆区与磷酸酶的结合,对T细胞的活化有负调节作用。
7、CD40L(CD154):
是TNF超家族成员,表达在CD4 细胞及部分CD8细胞上,以三聚体形式与B细胞上的CD40结合,是B细胞进行免疫应答的第二信号。
第三节 参与B细胞抗原识别与活化的CD分子
包括----与膜表面Ig共同形成的BCR复合物的CD79a-CD79b;具有辅助BCR功能的 CD19/CD21/CD81复合物;协同刺激信号受体CD80/CD86(B7-1/B7-2) 还有CD40、CD22等
1、Ig α/ Igβ(CD79a/CD79b)
通过二硫键组成异源性二聚体,表达在除浆细胞以外的各个阶段B细胞膜上,是B细胞的特征标志。 CD79a/CD79b以非共价键与mIg相连,组成BCR
复合物,与T细胞的CD3的作用十分相象,即介导由BCR途径的信号传导。
2、CD19/CD21/CD81复合物
是B细胞的重要标志(浆细胞除外),辅助BCR复合体激活B细胞。
CD21又称补体受体2(CR2),胞外为15-16个CCP结构域组成。(CCP:4个半胱氨酸,C1-4、C2-3间以二硫键形成的球状结构)。
CD21的第一、二个CCP是结合补体C3d的部位。Ag-Ab-C复合物一端通过Ag与BCR结合,另一端通过C与CD19/CD21/CD81复合物上的CD21结合,交连作用使CD21上的PTK发生磷酸化,导致CD19上酪氨酸磷酸化,增强B细胞的活化。CD81为四次跨膜肽链,配体不清。
3、CD80/CD86(B7-1/B7-2):
在胞外有同源性,胞质区则无。静止的单核细胞、DC上CD80表达低,CD86较高,细胞活化时都明显增加。CD28/CD152结合,为T细胞TCR-CD3活化途径提供协同刺激信号。
4、CD40:属TNFR-SF,膜外区有4个富含半胱氨酸的重复序列,广泛表达在各类APC及B细胞上(浆细胞除外),与T细胞上的CD40L结合,提供B细胞活化的第二信号。
5、CD22:B细胞活化中的抑制性受体。B细胞活化时表达增加。
第四节 免疫球蛋白Fc段受体
五类Ig的不同功能与其结构有关,可通过与细胞表面不同的的Fc受体结合,介导Ig的生理功能,或参与不同的病理损伤过程。
Fc受体包括FcγR、FcαR、FcεR。FcγR又分为 FcγRI、 FcγRII、 FcγRIII CD编号 为 FcαR CD编号 为 FcεR分为FcεRI、 FcεRII, FcεRII CD编号 为
1、FcγRI (CD 64): 表达的细胞有单核、巨噬细胞 、DC ;可结合Ab类别有IgG1、3(高) 效应 为ADCC、促吞噬、促细胞释放IL-1、6、TNF等因子。
2、FcγRII(CD32):又分为FcγRII-A、 FcγRII-B;分布广泛,除红细胞外,其他血细胞都表达;FcγRII-A属低亲和力受体,介导吞噬作用。
FcγRII-B介导免疫抑制。
3、FcγRIII(CD16):是IgG的低亲和力受体。有跨膜型(FcγRIII-A)和GPI连接(FcγRIII-B)两种型别。跨膜型表达在NK、巨噬细胞、肥大细胞。 GPI
连接的表达在中性粒细胞。 CD16 与FcεRIγ链或TCR-CD3连接,传递活化信号,介导ADCC吞噬。GPI:糖基磷脂酰肌醇
4、FcαR(CD89):分布在巨噬细胞、T、B细胞,是IgA的中亲和力抗体。介导吞噬、介质释放及ADCC。
5、 FcεRI :肽链组成方式αβγ2,βγ链上都有ITAM,是IgE的高亲和力受体,变应原与肥大细胞、嗜碱性粒细胞上的IgE- FcεRI 复合物结合, FcεRI发生交连,传递信号,介导I型变态反应。
6、FcεRII(CD23):凝集素样结构,并形成三聚体。CD23表达在B细胞、单核细胞,是IgE的低亲和力受体。CD23可被蛋白酶水解形成可溶性CD23,它依然保留有三聚体凝集素样结构,可以结合IgE。膜CD23调节B细胞分泌IgE的能力(降低),可溶性CD23增加IgE的分泌。
-
医学免疫学 重点总结
第1章免疫学概论1、免疫:是机体识别自己,排除异己过程中所产生的生物学效应的总和,正常情况下是维持内环境稳定的一种生理性防御功能。…
-
医学免疫学重要复习总结
第一章免疫学的基本内容1免疫的概念免疫是机体识别排除抗原性异物的生理反应2免疫的三个基本功能免疫防御对外来异体抗原物质的识别排除免…
-
医学免疫学知识总结
第三章抗原AntigenAg一基本概念1抗原AntigenAg是一类能刺激机体免疫系统使之产生特异性免疫应答诱导产生抗体及效应T淋…
-
医学免疫学重点总结
医学免疫学重点总结医学免疫学重点总结第一讲绪论1概念1免疫immunity即免除疫病和抵抗疾病的发生是机体识别自己排除异己非己过程…
-
医学免疫学总结
医学免疫学第一章免疫学概说一概念免疫生物在生存发展过程中所形成的识别自我与非己以及通过排斥非己而保护自我的现象二免疫力的构成一固有…
-
医学免疫学 重点总结
第1章免疫学概论1、免疫:是机体识别自己,排除异己过程中所产生的生物学效应的总和,正常情况下是维持内环境稳定的一种生理性防御功能。…
-
医学免疫学考试重点总结
一绪论免疫对自己或非己的识别并排除非己以保护体内环境稳定的一种生理反应免疫防御即抗感染免疫机体针对外来抗原如微生物及其毒素的免疫保…
-
医学免疫学重要复习总结
第一章免疫学的基本内容1免疫的概念免疫是机体识别排除抗原性异物的生理反应2免疫的三个基本功能免疫防御对外来异体抗原物质的识别排除免…
-
医学免疫学重点笔记(精华版)
医学免疫学一免疫的概念机体对自己或非己的识别应答过程中所产生的生物学效应的总和正常情况下是维持内环境稳定的一种生理性功能担负着机体…
-
电子科技大学生物技术专业免疫学重点总结
医学免疫学重点总结第一讲绪论1概念1免疫immunity即免除疫病和抵抗疾病的发生是机体识别自己排除异己非己过程中所产生的生物学效…
-
《医学免疫学》教学总结
《医学免疫学》教学总结基础医学部微免教研室自学校升为本科院校之后,学校加强了本科专业建设的投入,加大了人才培养模式的改革的力度,经…