大学生物之细胞总结
第一章
一、什么是细胞生物学?
是从细胞整体、超微结构和分子水平上研究细胞的结构和生命活动规律的科学。
二、细胞生物学的主要研究内容:
细胞核、染色体以及基因表达研究;生物膜与细胞器研究;细胞骨架体系研究;细胞增殖及其调控;细胞分化及其调控;细胞的衰老与凋亡;细胞的起源与进化;细胞工程。
三、细胞生物学发展简史:
细胞的发现;细胞学说的建立;细胞生物学学科的建立与发展。
四、当前细胞生物学研究中的三大基本问题
1)基因组在细胞内是如何在时间和空间上有序表达的?
2 )基因表达的产物,主要是结构蛋白与核算、脂质、多糖及其复合物,如何逐级组装成能行使生命活动的基本结构体系和细胞器?
3 )基因表达的产物,主要是大量活性因子与信号分子,是如何调节诸如细胞的增殖、分化、衰老与凋亡等细胞最重要的生命活动过程的?
五、1838年德国植物学家施莱登发表《植物发生论》,指出细胞是构成植物的基本单位。1839年德国动物学家施旺发表了<<关于动植物的结构和生长的一致性的显微研究>>论文,指出动植物是细胞的集合物。施旺和施莱登两人共同提出:一切植物、动物都有细胞组成,细胞是一切动植物的基本单位,这就是著名的“细胞学说”。
六、 “细胞学说”的基本内容:细胞是有机体,一切动植物都是由细胞发育而来,并由细胞和细胞产物所构成;每个细胞作为一个相对独立的单位,既有它“自己的”生命,又对与其它细胞共同组成的整体的生命有所助益;新的细胞可以通过老的细胞繁殖产生。
第二章
一、1、一切有机体都有细胞构成,细胞是构成有机体的基本单位.。2、细胞具有独立的、有序的自控代谢体系,细胞是代谢与功能的基本单位。3、细胞是有机体生长与发育的基础。4、细胞是遗传的基本单位,细胞具有遗传的全能性。
4、没有细胞就没有完整的生命。6、关于细胞概念的一些新思考(细胞是物质(结构)、能量与信息过程精巧结合的综合体;细胞是多层次、非线性与多层面的的复杂结构体系;细胞是高度有序的,具有自组装能力与自组织体系)
二、细胞的基本共性:1)所有的细胞都有相似的化学组成2)脂-蛋白体系的生物膜3)DNA-RNA的遗传装置4)蛋白质合成的机器-核糖体5)一分为二的分裂方式
三、最小最简单细胞——支原体
四、真核细胞的基本结构体系:1)以脂质及蛋白质成分为基础的生物膜结构体系2)以核酸与蛋白质为主要成分的遗传信息表达体系3)由蛋白质分子组装构成的细胞骨架体系
五、限制细胞体积大小的因素:1 体积同表面积的关系2 细胞核控制细胞质的活动3 细胞内物质交流运输与细胞体积的关系
第三章
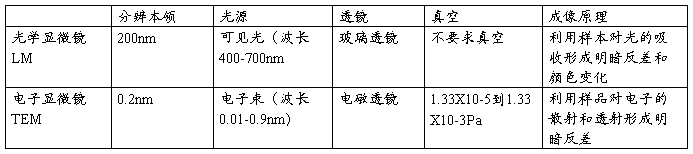
一、分辨率:是指区分开两个质点间的最小距离。 分辨率的高低取决于光源的波长λ,物镜镜口角α和介质折射率N。
D=0.61λ/(N.sin α /2)
二、1、动物细胞培养类型:原代细胞(primary culture cell)、传代细胞(subculture cell)、细胞系(cell line)有限细胞系(finite cell line)正常二倍体,接触抑制永生细胞系(infinite cell line)亚二倍体,接触抑制丧失细胞株(cell strain) 2、植物细胞:类型:单倍体细胞培养(花药培养)原生质体培养 (体细胞培养) 3、非细胞体系(cell-free system)
三、单克隆抗体(monoclone antibody):来自单个细胞克隆所分泌的抗体分子。
第四章
一、膜脂成分:磷脂: 分为二类: 甘油磷脂和鞘磷脂
主要特征:①具有一个极性头和两个非极性的尾(脂肪酸链)心磷脂除外);②脂肪酸碳链碳原子为偶数,多数碳链由16,18或20个组成;③饱和脂肪酸(如软脂酸)及不饱和脂肪
酸(如油酸); 糖脂:糖脂普遍存在于原核和真核细胞的质膜上(5%以下),神经细胞糖脂含量较高;ABO血型糖脂。胆固醇:胆固醇存在于真核细胞膜上(30%以下),调节膜的流动性,增加膜的稳定性以及降低水溶性物质的通透性。细菌质膜不含有胆固醇,但某些细菌的膜脂中含有甘油脂等中性脂类
二、脂质体的类型:(a)水溶液中的磷脂分子团;(b)球形脂质体;(c)平面脂质体膜;(d)于疾病治疗的脂质体的示意图
三、运动方式:沿膜平面的侧向运动;脂分子围绕轴心的自旋运动;脂分子尾部的摆动;双层脂分子之间的翻转运动。
四、膜蛋白类型:外在膜蛋白(extrinsic membrane protein);内在膜蛋白(intrinsic membrane protein);脂锚定膜蛋白(lipd membrane protein)
五、外在膜蛋白特性:外在膜蛋白为水溶性蛋白,靠离子键或其他较弱的键与膜表面的膜蛋白分子或膜质分子结合,因此只要改变溶液的离子强度甚至提高温度就可以从膜上分离下来,但膜结构并不被破坏。
脂锚定膜蛋白是通过与之共价相连的脂分子插入膜的脂双分子中,从而锚定在细胞质膜上。
六、去垢剂(detergent):是一端亲水另一端疏水的两性分子,是分离与研究膜蛋白的常用试剂。
离子型去垢剂如十二烷及磺酸钠(SDS)不仅可使细胞质膜崩解,还可与膜蛋白疏水部分结合从而使其与膜分离,而且还可以破坏蛋白质内部的非共价键,甚至改变其亲水部分的构象。由于SDS对蛋白质的作用较剧烈,引起蛋白质变性,因此在纯化膜蛋白时,特别是获得有生物活性的膜蛋白时,常采用非离子型去垢剂。
常用非离子型去垢剂如Triton-X-100, 非离子型去垢剂也可使细胞质膜崩解,但对蛋白质的作用比较温和。不同类型的非离子去垢剂,对不同种类的膜蛋白作用也有所不同,因此它不仅用于膜蛋白的分离与纯化,也用于除去细胞的膜系统,以便对细胞支架蛋白和其他蛋白进行研究。
七、生物膜基本特征:膜的流动性;膜的不对称性。
八、膜质流动性影响因素:
膜的流动性主要指脂分子的侧向运动,在很大程度由脂分子本身性质决定。一般来说,脂肪酸链越短,不饱和程度越高,脂膜的流动性越大,温度对膜质的流动性运动也有影响;在动物细胞中,胆固醇对膜的流动性也有双重调节作用。
***九、细胞质膜的基本功能:①为细胞的生命活动提供相对稳定的内环境;②选择性的物质运输,包括代谢底物的输入与代谢产物的排除,其中伴随着能量的传递;③提供细胞识别位点,并完成细胞内外信息跨膜转导;④为多种酶提供结合位点,使酶促反应高效而有序地进行;⑤介导细胞与细胞、细胞与胞外基质之间的连接;⑥参与形成具有不同功能的细胞表面特化结构;⑦膜蛋白的异常与某些遗传病、恶性肿瘤,甚至神经退行性疾病相关,很多膜蛋白可作为疾病治疗的药物靶标。
十、膜质的不对称性:指同一种膜质分子的脂双层中呈不均匀分布。糖脂的分布表现为完全不均匀。
膜蛋白的不对称性:指每种膜蛋白在膜质分子上都具有确定的方向性。
第五章
一、主动运输(active transport):一种需要消耗能量的物质跨膜运输过程。被运输底物与跨膜载体蛋白结合,通过载体蛋白构象改变,从而将底物逆着电化学梯度转运到膜的另一侧。
二、被动运输(passive transport):指通过简单扩散或协助扩散实现物质由高浓度向低浓度方向的跨膜转运。
三、简单扩散(simple diffusion):疏水的小分子或小的不带电荷的极性分子进行跨膜转运时,不需要细胞提供能量,也无需膜转运蛋白的协助,因此称为简单扩散。
四、协助扩散(facilitated diffusion):物质通过与特异性膜蛋白相互作用,高浓度向低浓度方向的跨膜转运。(这与简单扩散相同,两者都称为被动运输。)
五、协同转运(cotransport):是一类由Na+-K+泵与载体蛋白协同作用,靠间接消耗ATP所完成的主动运输方式。物质跨膜运动所需的直接动力来自膜两侧离子的电化学梯度,而维持这种离子电化学梯度则通过Na+-K+泵消耗ATP所实现。
动物细胞是利用膜两侧的Na+电化学梯度来实现的,而植物细胞和细菌常利用H+电化学梯度来实现的。
六、钠钾泵:具有ATP酶活性,因此又称Na+-K+ATPase。Na+-K+泵是由2个a亚基和2个β亚基组成的四聚体,β亚基是糖基化多肽,并不直接参与离子跨膜运输运动,但帮助在内质网新合成的a亚基进行折叠。
钠钾泵的作用:①维持细胞的渗透性,保持细胞体积;②维持低Na+高K+的细胞内环境;③维持细胞的静息电位。
地高辛、乌本苷等强心剂抑制其活性;Mg2+和少量膜脂有助提高于其活性。
七、钙离子泵:又称Ca+ATPase,是另一类P—型离子泵,分布在所有真核生物的质膜和某些细胞器膜上。Ca+泵由1000个氨基酸残基组成的多肽构成跨膜蛋白,与Na+-K+泵的a亚基同源,每一泵单位含有10个跨膜。a螺旋,其中3个螺旋与跨越脂双层的中央通道相连。作用:维持细胞内较低的钙离子浓度(胞内钙浓度10-7M,胞外10-3M)。
位置:质膜、内质网膜。
类型:P型离子泵,每分解一个ATP分子,泵出2个Ca2+。位于肌质网上的钙离子泵占肌质网膜蛋白质的90%。
钠钙交换器(Na+-Ca2+ exchanger),属于反向协同运输体系,通过钠钙交换来转运钙离子。
八、V-type:广泛存在于动物细胞内体、溶酶体膜,破骨细胞和某些肾小管细胞的质膜以及植物、酵母和其他真菌细胞液泡膜上,又称膜泡质子泵。
九、F-type:存在于细菌质膜、线粒体内膜和叶绿体类囊体膜上。利用质子动力势合成ATP,即ATP合酶
十、ATP驱动泵类型:P-型离子泵(钠钾泵、钙泵)、V-型质子泵、F-型质子泵、ABC超家族。
第六章
一、氧化磷酸化的偶联机制—化学渗透假说:◆1961年英化学家Mitchell提出,获1978年诺贝尔化学奖;◆化学渗透假说内容:电子传递链各组分在线粒体内膜中不对称分布,当高能电子沿其传递时,所释放的能量将H+从基质泵到膜间隙,形成H+电化学梯度。在这个梯度驱使下,H+穿过ATP合成酶回到基质,同时合成ATP,电化学梯度中蕴藏的能量储存到ATP高能磷酸键。◆支持化学渗透假说具有实验证据,该实验表明:·质子动力势(proton motive force) 乃ATP合成的动力·膜应具有完整性·电子传递与ATP合成是两件相关而又不同的事件
◆ ATP形成成机制:ADP转变为ATP是一个磷酸化的过程。ADP磷酸化有两个途径①底物水平磷酸化,由相关的酶将底物分子上的磷酸集团直接转移到ADP分子上,生成ATP。②氧化磷酸化,只在呼吸链上与电子传递相耦联的ADP被磷酸化形成ATP酶促过程。是需氧细胞的生命活动主要能量来源,是ATP生成主要途径。
二、叶绿体的功能—光合作用 (photosynthesis) Photosynthesis:
三大步骤:原初反应、电子传递和光合磷酸化、碳同化。(1)光反应(Light Reaction)——原初反应、电子传递和光合磷酸化(2)暗反应(Dark Reaction)——碳固定反应●光反应 在类囊体膜上发生,将光能转换为化学能(ATP、NADPH)●暗反应 (碳固定) 在叶绿体基质中进行,利用ATP和NADPH使CO2还原为糖类●光合作用与有氧呼吸的关系图 ;光合磷酸化与氧化磷酸化比较
三、光反应:在类囊体膜上由光引起的光化学反应,通过叶绿素等光合色素分子吸收、传递光能,水光解,并将光能转换为电能(生成高能电子),进而通过电子传递与光合磷酸化将电能转换为活跃化学能, 形成ATP和NADPH并放出 O2 的过程。包括原初反应、电子传递和光合磷酸化。◆原初反应(primary reaction)捕光(天线)色素+反应中心= 光合作用单位;反应中心=1中心色素+1原初电子供体D+1原初电子受体A
·光能的吸收、传递与转换,形成高能电子
◆电子传递与光合磷酸化
四、暗反应(碳固定) 利用光反应产生的ATP 和NADPH,使CO2还原为糖类等有机物,即将活跃的化学能最后转换为稳定的化学能,积存于有机物中。这一过程不直接需要光(在叶绿体基质中进行)。◆卡尔文循环(Calvin cycle)(C3途径)◆C4途径或 Hatch-Slack循环(C4植物:甘蔗、玉米等)
◆景天科酸代谢途径(CAM途径,与C4途径相似,只是时间空间上不同;CAM植物:景天、落地生跟等)
第七章
一、过氧化物酶体与溶酶体的区别:
1)过氧化物酶体和初级溶酶体的形态与大小类似,但过氧化物酶体中的尿酸氧化酶等常形成晶格状结构,可作为电镜下识别的主要特征。2)通过离心可分离过氧化物酶体和溶酶体 3)过氧化物酶体和溶酶体的差别
二、溶酶体酶甘露糖残基的磷酸化先后有两种酶催化:一种是N-乙酰葡糖胺磷酸转化酶,另一种是磷酸葡糖苷酶。
三、信号肽(signal sequence):位于蛋白质的N端,一般有16-26个氨基酸残基,其中包括疏水核心区、信号肽的C端和N端3部分。
四、信号假说(signal hypothesis):即分泌性蛋白N端序列作为信号肽,指导分泌性蛋白到内质网膜上合成,然后在信号肽引导下蛋白质边合成边通过易位子蛋白复合体进入内质网腔,在蛋白质合成结束之前信号肽被切除。
五、三种不同的膜泡运输方式及作用:①COPII有被小泡的组装与运输→COPII有被小泡介导细胞内顺向运输从内质网到高尔基体的物质运输; ② COPI有被小泡的组装与运输→COPI有被小泡介导细胞内逆向运输从顺面高尔基体网状区到内质网膜泡转运,包括再循环的膜脂双层、某些蛋白质如v-SNAREs和回收错误分选的内质网逃逸蛋白返回内质网。③网格蛋白有被小泡的组装与运输→网格蛋白有被小泡介导蛋白质从高尔基体TGN向质膜、胞内体、溶酶体或植物液泡运输;
六、内质网标致酶:蛋白二硫键异构酶,它附着在内置网膜腔面上,可切断二硫键,形成自由能最低的蛋白构象,以帮助新合成的蛋白重新形成二硫键并处于正确折叠状态。
七、过氧化物酶体(peroxisome)又叫微体(microbody),是由单层膜围绕的内含一种或几种氧化酶类的细胞器。两种酶是:第一种是依赖于黄素(FAD)的氧化酶,作用是将底物氧化形成H2O2;第二种是过氧化氢酶,作用是将H2O2分解形成水和氧气。
**八、溶酶体酶的合成、修饰及分选过程
溶酶体的发生:溶酶体酶在粗糙内质网上合成并经N-连接的糖基化修饰,然后转至高尔基体,在高尔基体的顺面膜囊中寡糖链上的甘露糖残基被磷酸化形成M6P,在高尔基体的反面膜囊和TGN膜上存在M6P的受体,这样溶酶体与其他蛋白质区分开来并得以浓缩,最后以出芽的方式转运到溶酶体中。
分选途径多样化:依赖于M6P 的分选途径的效率不高,部分溶酶体酶通过运输小泡直接分泌到细胞外;在细胞质膜上也存在依赖
于钙离子的M6P受体,同样可与胞外的溶酶体酶结合,通过受体介导的内吞作用,将酶送至前溶酶体中,M6P受体返回细胞质膜,反复使用。还存在不依赖于M6P的分选途径(如酸性磷酸酶、分泌溶酶体的perforin和granzyme)
第八章
一、细胞通讯(cell communication):是指一个细胞发出的信息通过介质传递到另一个细胞并与靶细胞相应的受体相互作用,然后通过细胞信号转导产生胞内一系列生理生化变化,最终表现为细胞整体的生物学效应过程。
二、细胞通讯方式:①细胞通过分泌化学信号进行通讯(内分泌(endocrine)、旁分泌(paracrine)、自分泌(autocrine)、化学突触(chemical synapse))②细胞间接触性依赖的通讯,指细胞间直接接触,通过与质膜结合的信号分子影响其他细胞。③动物相邻细胞间形成间隙连接以及植物细胞间通过胞间连丝使细胞间相互沟通,通过交换小分子来实现代谢耦联或电耦联。
三、各种化学信号根据溶解性分为亲脂性和亲水性两类。
四、受体(receptor):是一种能够识别和选择性结合某种配体的大分子,绝大多数已经鉴定的受体都是蛋白质且多为糖蛋白,少数受体为糖脂,有的受体为糖蛋白和糖脂组成的复合物。
五、细胞表面受体包括:离子通道耦联受体;G蛋白耦联受体;酶连受体。
六、NO作用于邻近细胞。
NO在血管内皮细胞和神经细胞中生成,由一氧化氮合酶(NOS)催化,以L精氨酸为底物,NADPH为电子供体,生成NO和L瓜氨酸。
NO的作用机理:乙酰胆碱→血管内皮→Ca2+浓度升高→一氧化氮合酶→NO→平滑肌细胞→鸟苷酸环化酶→cGMP→血管平滑肌细胞的Ca2+离子浓度下降→平滑肌舒张→血管扩张、血流通畅。
硝酸甘油治疗心绞痛具有百年的历史,其作机理是在体内转化为NO。
NO作为受体信号分子引起血管平滑肌舒张;NO是一种自由基性质的气体,具脂溶性,可快速扩散透过细胞质膜,到达邻近靶细胞发挥作用。
第九章
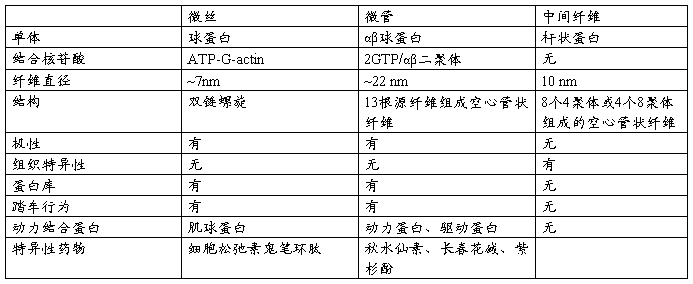
一、细胞骨架(Cytoskeleton)包括:微丝(microfilament,MF)微管(microtubule,MT)、中间丝(intermediate filament ,IF)
二、分子马达(molecular motor):主要指依赖微管的驱动蛋白、动力蛋白和依赖于微丝的肌球蛋这三类蛋白质的超级家族的成员。
三、微观组织中心(microtubule organizing center, MTOC):在活细胞内,能够起始微管的成核作用,并使之延伸的细胞结构称为微观组织中心。
四、分子伴侣(chaperone,molecular chaperon):一种与其他多肽或蛋白质结合的蛋白质,以防止蛋白质错误折叠、变性或聚集沉淀,对蛋白质的正确折叠、组装以及跨膜转运有意义。
五、马达蛋白(motor protein):利用ATP水解释放的能量驱动自身沿微管或微丝定向运动的蛋白,如驱动蛋白、动力蛋白和肌球蛋白。
第十章
一、核孔复合体功能:核质交换的双向选择性亲水通道。
从功能上讲,核孔复合体可以看作是一种特殊的跨膜运输蛋白复合体,并且是一个双功能、双向性的亲水性核质交换通道。双功能表现在它两种运输方式:被动扩散与主动运输。双向性表现在即介导蛋白质的入核转运,又介导RNA、核糖核蛋白颗粒的出核转运。
二、间期染色质按其形态特征、活性状态和染色性能分为:常染色质和异染色质。
常染色质(euchromatin):是指间期细胞核内染色质纤维折叠压缩程度低,相对处于延展状态,用碱性染料染色时着色浅的那些染色质。
异染色质(heterochromatin):是指间期核中,染色质纤维折叠压缩程度高,相对处于聚缩状态,用碱性染料染色时着色深的那些染色质。
三、结构异染色质特征:在中期染色体上多定位于着丝粒区、端粒、次缢痕及染色体臂的某些节段;由相对简单、高度重复的DNA序列构成,如卫星DNA;具有显著遗传惰性,不转录也不编码蛋白质;在复制行为上与常染色质相比表现为晚复制早聚缩;占据较大部分核DNA,在功能上参与染色质高级结构的形成,导致染色质区间性,作为核DNA的转座元件,引起遗传变异。
四、DNA结合蛋白的结构模式:a螺旋-转角-a螺旋模式(helix-turn-helix motif);锌指模式(zinc finger motif);亮氨酸拉链模式(leucine zipper motif,ZIP);螺旋-环-螺旋结构模式(helix-loop-helix motif,HLH);HMG框结构模式(HMG-box motif)
***五、核小体结构要点:1)每个核小体单位包括200bp左右的DNA超螺旋和一个组蛋白八聚体以及一个分子的组蛋白H1;2)组蛋白八聚体构成核小体的盘状核心颗粒,相对分子质量100000,由4个异二聚体组成,包括两个H2A.H2B和两个H3.H4;3)146bp的DNA分子超螺旋盘绕组蛋白八聚体1.75圈;4)两个相邻核小体之间以连接DNA相连,典型长度60bp,不同物种变化值为0-80bp不等;5)组蛋白与DNA之间相互作用主要是结构性的,基本不依赖于核苷酸的特异序列;6)核小体沿DNA的定位受不同因素的影响,
六、次缢痕(secondary constriction):在染色体上其他的浅染缢缩部位称次缢痕。它的数目、位置和大小是某些染色体所特有的形态特征,可以作为鉴定染色体的标记。
七、 核仁组织区(nucleolar organizing region,NOR):位于染色体的次缢痕部位,并非所有次缢痕都是NOR。染色体NOR是rRNA基因所在部位,与同期细胞核仁形成相关。
八、 端粒(telomere):位于染色体末端的重复序列,对染色体结构稳定、末端复制等有重要作用。端粒常在每条染色体末端形成一顶“帽子”结构
九、端粒酶(telomerase):含有RNA反转录酶,能以自身RNA为模板,对DNA端粒序列进行延长而解决线性染色体末端复制问题。
十、真核细胞的核仁具有重要功能:它是rRNA合成、加工和核糖体亚单位的组装场所。
十一、纤维中心(fibrillar centers,FC):是包埋在颗粒组分内部一个或几个浅染的低电子密度的圆形结构。它存在于rDNA 、RNA聚合酶I和结合的转录因子。
十二、致密纤维组分(dense fibrillar component,DFC):它是核仁超微结构中电子密度最高的部分,呈环形或半月形包围FC,由致密的纤维构成,通常见不到颗粒。
十三、颗粒组分(granular component,GC):是核仁的主要结构,由直径15-20nm的核糖核蛋白颗粒构成,可被蛋白酶和RNase消化,这些颗粒是正在加工、成熟的核糖体亚单位前体颗粒,间期核中核仁的大小差异主要是由颗粒组分数量的差异造成的。
十四、核仁组分包括:核仁相随染色质、核仁内染色质、核仁周边染色质。
十五、真核生物核糖体含有4种rRNA:即5.8S rRNA、18S rRNA、28S rRNA、及5S rRNA。即前3种基因组成一个转录单位。与为蛋白质编码的mRNA不同,rRNA分子是rRNA基因的最终产物。
十六、DNA结合蛋白包括:组蛋白(histone),与DNA结合但没有序列特异性;非组蛋白(nonhistone),与特定DNA序列或组蛋白相结合。
十七、核定位序列或核定位信号:存在于亲核蛋白内的,具有定向、定位作用的一些氨基酸序列片段,富含碱性氨基酸残基,如Lys、Arg,还常常含有Pro,并且在指导亲核蛋白完成核输入后并不被切除。
第十二章
一、 周期蛋白(cyclin protein):在海胆卵细胞中存在两种特殊蛋白质,这两种蛋白质的含量随细胞周期进程变化而变化,一般在细胞间期内积累,在细胞分裂期内消失。在下一个细胞周期中又重复这一消长现象,因此将这两种蛋白质命名为周期蛋白。
二、 周期蛋白结构特点:① 均含有一段保守的氨基酸序列,称为周期蛋白框;② 介导Cyclin与CDK 结合;③不同的cyclin在细胞周期中表达的时期不同,并与不同的CDK结合,调节不同的CDK酶活性。
三、 CDK激酶 (cyclin-dependent kinase) 和CDK激酶抑制剂结构特点: ①都含有一段类似的氨基酸序列(PSTAIRE)序列。②都可以与cyclin结合,并将其作为调节亚基表现激酶活性。③不同的CDK结合的cyclin不同。
CDK由cdc基因编码产生,具蛋白激酶活性,是整个细胞周期的关键调节者,它在细胞增殖周期中的活性变化受细胞各种信号的精确调节,使细胞的增殖分化与环境和发育相适应
CDK活性调节:周期蛋白结合活化;修饰性调控因子(磷酸化等);CDK激酶抑制物(CDKI) Cdc2激酶被命名为CDK1,可将特定蛋白磷酸化,促进细胞周期运行,因此又称做细胞周期引擎。
CDK分子中还存在一些重要位点,这些位点的磷酸化与去磷酸化,对CDK活性的调节起重要作用。
四、 CDK1激酶可以使多种蛋白质磷酸化,其中包括组蛋白H1、核纤层蛋白A、B、C、核仁蛋白和No.38、p60 c-src、 C-abl等,组蛋白H1磷酸化,促使染色质凝结;核纤层蛋白磷酸化,促使核纤维层解聚;核仁蛋白酶磷酸化,促使核仁解体;p60 c-src蛋白磷酸化,促使细胞骨架重排;C-abl蛋白磷酸化,促使形态调整。
第十三章
一、细胞坏死与细胞凋亡区别:对于动物细胞,细胞坏死是区别于细胞凋亡的另一种典型细胞死亡方式。长期以来细胞坏死被认为是一种被动的死亡方式,当细胞受到意外损伤,如极端的物理、化学因素或严重的病理性刺激的情况下细胞坏死才会发生。此时细胞内ATP浓度已下降到无法维持细胞存活水平。细胞坏死时,细胞质出现空泡,细胞质膜破损,细胞内含物,包括膨大或破碎的细胞器以及染色质片段释放到胞外,引起周围组织炎症反应。与细胞凋亡不同,细胞坏死过程中染色质不发生凝集,也不产生有规律的200bp的DNA降解片段,而是被随机降解,琼脂糖凝胶电泳时呈现弥散性分布,俗称“拖尾”现象。
第十四章
一、癌细胞的基本特征:细胞生长与分裂失去调控;具有浸 和扩散性;细胞间相互作用改变;mRNA的表达谱及蛋白质表达谱或蛋白活性改变;体外培养的恶性转化细胞的特征。
第十五章
一、细胞连接(cell junction):在细胞质膜的特化区域,通过膜蛋白、细胞支架蛋白或者胞外基质间的结构。
二、细胞连接的类型:①封闭连接(occluding junction)将相邻的上皮细胞的质膜紧密的连接在一起,阻止溶液中的小分子沿细胞间隙从细胞一侧渗透到另一侧。紧密连接是典型代表。②锚定连接(anchoring junction)通过细胞质膜蛋白及细胞骨架系统将相邻细胞或细胞与胞外基质间连接起来。根据直接参与细胞连接的细胞骨架纤维种类不同又分为中间丝相关的锚定连接和肌动蛋白丝相关的锚定连接。前者包括桥粒(desmosome)和半桥粒(hemidesmosome)后者包括黏合带(adhesion belt)和黏合斑(focal adhesion)③通讯连接(communication junction)介导相邻细胞间的物质转运、化学或电信号的传递,包括间隙连接(gap junction)神经元间的化学突触(chemical synapse)和植物细胞间的胞间连丝.
-
细胞生物学总结(复习重点)——1-2.绪论与简介
1、细胞生物学cellbiology:是研究细胞基本生命活动规律的科学,是在显微、亚显微和分子水平上,以研究细胞结构与功能,细胞增…
-
细胞生物学考试总结
一名词解释1细胞生物学研究细胞的结构功能和生命活动基本规律的科学2细胞培养把机体内的组织取出后经过分散机械方法或酶消化为单个细胞在…
-
细胞生物学重点总结
第一章细胞生物学绪论1什么是细胞什么是细胞生物学细胞是细胞宇宙有机界一个非常重要的层次它一方面是由质膜包围的相对独立的功能单位能够…
-
细胞生物学第三版 重点总结
第一章绪论一细胞生物学的主要研究内容一细胞核染色体以及基因表达的研究二生物膜与细胞器的研究三细胞骨架体系的研究四细胞增值及其调控五…
-
细胞生物学总结+
细胞生物学总结20xx级药学第一章二章1什么是细胞什么是细胞生物学细胞是生物的形态结构和生命活动的基本单位细胞生物学以细胞为研究对…
-
20xx年上海交通大学生物化学考研资料参考书目历年真题笔记解析
生物化学831参考书目:《生物化学》第三版沈同、王镜岩主编高等教育出版社真题系列(纸质版):20xx生物化学试题20xx生物化学试…
-
写给20xx华南理工大学考研复习生物化学的一点心得体会
写给20xx华南理工大学考研复习生物化学的一点心得体会许多20xx年报考华南理工大学食品的同学都选择了生物化学这门专业课,但是往往…
-
20xx中山大学生物化学复习方法 之往届复习心得
考完了,也终于有时间有机会为那艰苦的岁月记下点东西了~~考生物学是一种挑战。我比较喜欢挑战,于是选择了生物学专业。第一眼见到王镜岩…
-
大学生化复习总结
蛋白质分子组成组成单位氨基酸结构特点氨基羧基L构型分类碱性HisArgLys酸性GluAsp中性理化性质水溶性pI紫外吸收含氮量1…
-
大学生物化学简答题总结(蛋白质)免费
第二章蛋白质蛋白质分类1根据蛋白质分子的外形可以将其分作3类球状蛋白质分子形状接近球形水溶性较好种类很多可行使多种多样的生物学功能…
-
细胞生物学考试重点总结
名词解释1Hayflick界限:正常的体外培养细胞的寿命不是无限的,而只能进行有限次数的增值约50次2细胞连接celljuncti…