光与光合作用论文
《光与光合作用》教学设计与反思(2课时)
引言:“学习科学是学生们要亲自动手做而不是要别人做给他们看的事情”。运用现代教育技术,充分发挥学生的主体作用。即教师创设教学情景,留给学生更多的时空,让学生尽可能多地参与知识的产生、发展的过程,变被动接受知识为主动发现知识、掌握知识,从而达到培养学生学习能力和创造能力的目标。
一、教材分析
本节课为高中必修一《分子与细胞》人教版第一册第五章“细胞的能量供应和利用”中的第四节的内容。本册教材第五章介绍了细胞的能量供应和利用,本节课是对细胞的能量来源进行探讨,为以后进行教学知识打下基础具有承上启下的作用。因此,使学生牢固掌握光合作用这一部分知识非常重要。光合作用部分共有五个问题:光合作用的概念、叶绿体中的色素、光合作用的过程、实质、意义。重点是叶绿体中的色素、光合作用的过程和意义,难点是光合作用的过程。本节内容的学习会对学生以后学习有关能量来源问题和实验设计有很好的促进作用。
本节课进行光合作用过程中氧气来源的实验设计,以及提取吸收光能的色素的实验操作,采取小组合作的方式,力图使学生在小组内讨论和组间交流及全班交流的过程中,学会合作,学会倾听,学会交流, 在此过程中巩固实验设计的基本方法,并在后续的课堂教学中完成动手操作,根据在实验过程中遇到的新问题,重新完善设计方案。
二、学习者特征分析
本节课的学习者特征分析主要是根据教师平时对学生的了解而做出的:
(1)学生是临沧市凤庆县第二中学高中一年级的学生;
(2)学生活泼好动,好表现,喜欢交流;
(3)学生已经知道植物能够进行光合作用,能够产生氧气。但光合作用的反应过程及其实质,光合作用释放的氧气具体来自哪里等知识了解不多;
(4)学生在以前运用实验法进行过简单的实验设计,但方案的科学性、可操作性都不是很强。
三、教学目标设计:
知识目标:
1.了解光合作用的概念、实质
2.掌握叶绿体中色素的种类和作用
3.掌握光合作用的过程和意义
技能目标:
1.通过对光合作用发现史的学习,了解科学家的科学思维方法
2.通过实验(含设计),培养学生初步的研究能力;
3.在有关实验、资料分析、思考与讨论、探究等的问题讨论中,运用语言表达的能力以及分享信息的能力。
4.利用精心设计的光合作用过程的多媒体动画,培养学生的比较、归纳、综合能力;
5.利用巧妙设计的练习题,提高学生分析、解决问题的能力。
情感目标:
结合教学内容,对学生进行科学史教育,树立辩证唯物主义的世界观、科学观,增强环保意识。
四、教学策略选择与设计
(1)情境激趣策略:通过动手提取叶绿体中吸收光能的色素的过程,加强学生的动手能力,调动学生的积极性;通过播放光合作用的动画演示,提出问题,激发学生的学习兴趣。
(2)以问题解决为主教学策略:通过观察,引发学生提出关于光合作用实质以及光合作用释放氧气的来源的疑惑,通过假设和实验方案的设计,得出科学性的结论,解决问题和疑惑。
(3)按照以学生为主体、教师为主导、训练为主线的“三主”教学原则,在教学过程中,采用教师讲授与学生自学相结合,教师的启发与学生的分析、讨论、归纳相结合,师生共同探究的方法,突出重点、突破难点。
五、学法指导:
探究学习法:对于光合作用的发现史,叶绿体中色素的提取、分离及作用,光合作用产物氧气的原子来源等采用科学探究的学习方法。
观察对比法:对光合作用的过程,采用先观察CAI 课件,再比较该过程中光反应与暗反应的区别和联系。
归纳教学法:由光合作用过程归纳出它的实质和意义。
六、教学资源与工具设计:
计算机、CAI课件(展示光合作用的发现史、色素的提取和分离的实验录象、色素的吸收光谱、光合作用的过程等)、实物展示平台(展示学生的实验结果)、实验材料等。
七、教学重难点:
重点:绿叶中色素的种类和作用;光合作用的发现和研究历史;光合作用的光反映、暗反应过程以及相互关系;影响 光合作用强度的环境因素。
难点:光反映和暗反应的过程;探究影响光合作用强度的环境因素。
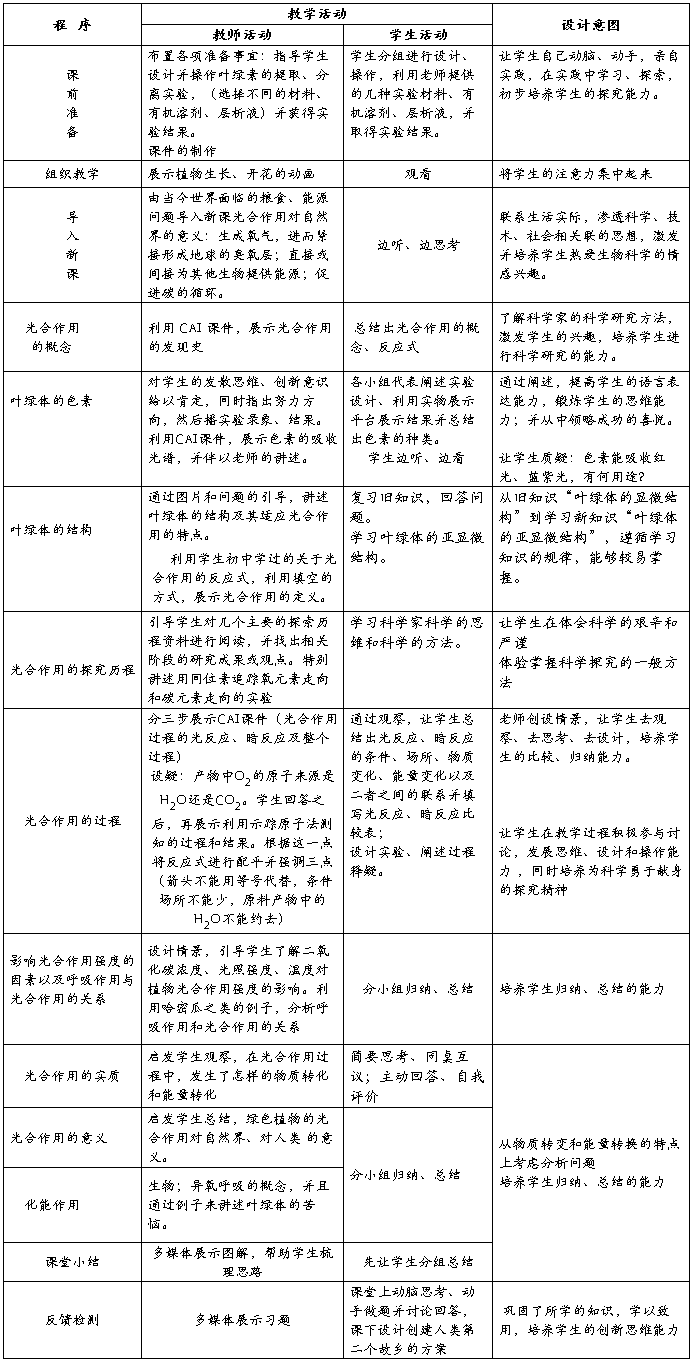
八、教学过程
九、教学评价设计
1.针对性练习
让学生思考完成具有针对性的练习,进行知识巩固和教学效果反馈,及时纠正错误的理解和片面的认识。
2.布置作业、及时巩固
在作业的布置中,我严格遵循“重质量、轻负担”的指导思想。题目的设定主要是为了帮助学生及时纠正原有的对知识的错误理解或片面认识,培养学生的解题技巧和技能。同时想借助该题培养学生对问题的科学的思维方法和探究的精神。
十、板书设计
第三节 能量之源——光与光合作
一、光合作用的概念
二、叶绿体中的色素
1. 叶绿体的色素
2. 叶绿体的结构
三、光合作用的过程、实质、意义
1.过程
2.实质
3.意义
十一、教学反思
1.教学过程中始终追求“学生为主体、教师为主导”新课程的教育体系要求教学过程要以学生的发展为本,提出“学生是教学的主体”。因此,我在备课、做课件时博取众长,吸收大家的精华,上课时针对教学过程,特别遵循学生从感性到理性的认知规律,充分考虑学生的情感、认知心理,循序渐进。在教学实施中,通过相应的现象或情境引入,然后让学生自己观察实验现象,讨论实验的相关设计和目的;然后通过讨论由学生自己去归纳,做出结论,充分发挥了学生的主观能动性,使学生进入到一种主动学习的状态。同时,我注意观察学生的学习状态和反馈,紧紧抓住学生的思维火花,及时引导、设问,激发其强烈的求知欲,成为了教学活动的组织者。
2.课题特点和教学设计有关光合作用的知识是整个高中教学的难点和重点。
我希望学生通过本节课的学习,达到“掌握科学实验探究的一般原则,重点是对照实验原则和单因子变量原则”的目标,为以后生物科学实验知识的学习、科学素养的培养做好铺垫和扫除一些障碍。所以我在教材处理上,一是以图片引出对色素、叶绿体结构的相关内容的学习,并结合彩图进行讲解,引起学生兴趣,利于学生掌握,并从中体会生物体结构与功能相适应的特点;二是按时间线索对有关光合作用的经典实验用PPT课件进行了图片、动画的补充学习,沿着科学家的脚迹进行了光合作用的发现探究之旅,让学生在体会科学的艰辛和严谨的同时,体验掌握了科学探究的一般方法;并点明科学发现是一场永不结束的接力赛,等着我们去接上下一棒,并奋力向前。
3.及时反馈对于重点知识点的学习,我注意在学后及时反馈,以便了解学生掌握和发现问题。如色素及叶绿体结构知识的学习后,我都结合了相关的一道选择题进行反馈;对光合作用的发现探究之旅学习后,结合了一个空的表格,让学生填表。
4.需改进之处 本节课也有不少失误的地方,最重要的是:由于本节课内容较多,科学实验也多,跨度大,学生又没有科学探究实验设计的基础,在学生讨论、师生互动的过程中,我在时间把握上没能做到恰到好处,导致后面内容的教学显得非常紧迫。
第二篇:干旱胁迫下镜面掌光合作用的动态变化研究_园林设计毕业论文
东北农业大学学士学位论文
干旱胁迫下镜面掌光合作用的动态变化研究
Bachelor Degree Paper of Northeast Agricultural University No: A02090043
The dynamic changes of photophysiological characters
of the mirror palm under drought stress
Name:
Supervisor:
Department:
Major:
Field:
Liu Fenfen Prof. Chen Yajun Horticulture Garden Ornamental plant
Northeast Agriculture University
China ·Harbin
June 2013
干旱胁迫下镜面掌光合作用的动态变化研究
摘 要 镜面掌是最普遍的室内观叶植物之一,其具有植株低矮,叶形美丽,色泽青绿,繁殖力强,生命力旺盛,养护成本低等优点而受人们喜爱。本研究通过温室盆栽试验方法,研究了观叶植物镜面掌Pilea peperomioides Diels在不同梯度干旱胁迫及复水后的光合生理反应,探索在干旱胁迫下镜面掌的光合生理指标的变化,分析干旱胁迫程度对各光合生理指标的影响,为镜面掌抗旱生理的研究提供理论依据和技术支持。本研究主要结果如下:
在干旱胁迫下,镜面掌的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)四项光合生理指标均有显著变化,随胁迫程度的加重净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)均有显著降低,胞间CO2浓度(Ci)呈现先降低、但最终上升而累积的现象,说明干旱胁迫可以显著影响植物的光合作用,且干旱胁迫的天数越多镜面掌光合能力下降的越明显,即干旱胁迫程度越大对植物的影响也越大。在对镜面掌干旱胁迫后进行复水处理,净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)四项指标均有所增加,干旱胁迫天数越少的,其恢复能力也越强,但是都显著低于对照的水平。
关键词:镜面掌;干旱胁迫;光合生理
I
干旱胁迫下镜面掌光合作用的动态变化研究
Abstract Under drought stress, the mirror palm return of net photosynthetic rate (Pn), stomatal conductance (Gs), transpiration rate (Tr), intercellular CO2 concentration (Ci) photosynthetic physiological indexes of four significant changes, with the stress degree increased net photosynthetic rate (Pn), stomatal conductance (Gs) and transpiration rate were significantly lower (Tr), intercellular CO2 concentration (Ci) to reduce, increase accumulation phenomenon but in the end,Shows that drought stress can significantly affect plant photosynthesis, and the more the number of days drought stress mirror palm photosynthetic capacity decreased, the more obvious, namely, the influence of the greater the degree of drought stress on plant. After palm of drought stress to water treatment, net photosynthetic rate (Pn), stomatal conductance (Gs), transpiration rate (Tr),intercellular CO2 concentration (Ci) four indicators were increased, the less drought stress days, its recovery ability is stronger, but were significantly lower than control.
Key words: mirror palm; drought stress; photophysiological characters
II
干旱胁迫下镜面掌光合作用的动态变化研究
目录
摘 要 ...................................................................................................................................... I 1 前言...................................................................................................................................... - 1 -
1.1 研究目的和意义 ........................................................................................................ - 1 -
1.2植物对干旱胁迫的响应 ............................................................................................. - 1 -
1.3干旱胁迫对光合作用的影响 ...................................................................................... - 2 - 2 试验材料与方法 .................................................................................................................. - 4 -
2.1试验材料 .................................................................................................................... - 4 -
2.2试验设计 .................................................................................................................... - 4 -
2.3试验内容与方法 ......................................................................................................... - 4 -
2.3.1光合生理指标动态变化的测定 ....................................................................... - 4 -
2.4数据处理与分析 ......................................................................................................... - 5 - 3 结果与分析 .......................................................................................................................... - 6 -
3.1干旱胁迫条件下镜面掌叶片净光合速率(Pn)的变化 ............................................ - 6 -
3.2干旱胁迫条件下镜面掌叶片蒸腾速率(Tr)的变化 ................................................ - 6 -
3.3干旱胁迫条件下镜面掌片气孔导度(Gs)的变化 ................................................... - 7 -
3.4干旱胁迫条件下镜面掌叶片气孔阻力(Rs)的变化 ............................................... - 8 -
3.5干旱胁迫条件下镜面掌叶片胞间CO2浓度(Ci)的变化 ........................................ - 9 - 4 讨论.................................................................................................................................... - 10 -
4.1干旱胁迫下气孔结构指标与气孔导度(Gs)、蒸腾速率(Tr)及光合速率(Pn)的关系.................................................................................................................................... - 10 -
4.2干旱胁迫对镜面掌叶片光合特性的影响分析 ......................................................... - 10 - 5 结论.................................................................................................................................... - 12 - 参考文献 ................................................................................................................................ - 13 - 致谢 ....................................................................................................................................... - 15 -
III
干旱胁迫下镜面掌光合作用的动态变化研究
1 前言
镜面掌(Pilea peperomioides Diels)为荨麻科冷水花属多年生肉质草本植物,是一种罕见的耐寒喜阴植物[1],只要达到可看一般报纸的光亮即可生长[2],在阳光充足的温室内也生长良好,生长适宜的温度为15℃左右。适于在比较湿润排水良好的泥炭土上生长。镜面掌叶片深绿色,有光泽,是我国主要的室内观叶花卉[3],野生群体较少,在我国大多为人工室内栽培[4],在云南及华南以外其他地区多作温室盆栽。在我们人类生活的环境中,水分是不可或缺的,它也是植物生长的主要限制因子之一[5],植物的光合作用也离不开水分,因此,研究干旱胁迫下镜面掌光合作用的变化对镜面掌的抗旱机理及抗旱品种的选育具有重要意义
[6]。本研究采用温室盆栽镜面掌为试验材料,研究其在干旱胁迫下光合生理各项指标的变化。
1.1 研究目的和意义
随着生活水平的提高,人们利用绿色植物进行居室绿化及装饰已成为一种时尚[7]。最近,美国航空航天局的科学家们发现,室内观叶植物不但起到柔化空间美化环境等效果,调节室内的温度和湿度,而且可以吸收空气中的有害气体如甲醛、甲苯、二氧化硫等,起到净化室内空气的作用[8]。此外,室内观叶植物枝叶有滞留尘埃、吸收生活废气、减轻噪音,散发宜人香气等作用,因此室内养植观叶植物对营造良好生活环境具有重要的作用[9]。在社会经济不断发展的今天,人们对工作、生活环境的要求越来越高,室内观叶植物的应用也越来越被人们所接受,并逐渐受到人们的重视,在各类公共区域和居室环境被大量使用,尤其是经济较发达的长三角地区,各种新建和高档的厅、堂、馆、所、茶楼、酒店、银行、商厦乃至家庭住所布置一些清新、自然的室内观叶植物成为一种必不可少的装饰。室内观叶植物几乎能周年生长,深受人们的喜爱,在家庭、宾馆、大厦、办公室和餐厅等公共场所,都能见到他们的身影[10]。在室内观叶植物中,镜面掌姿态细软,叶片饱满充盈,色泽圆润,生长旺盛,因此成为室内装饰植物的首要选择。干旱胁迫是植物生长过程中常见的逆境之一,能够影响植物的正常生长[11]。光合作用是植物体内重要的代谢过程,绿色植物通过光合作用将太阳能转换为化学能,并利用它把二氧化碳和水等无机物合成有机物,同时放出氧气。绿色植物的光合作用不但与植物本身的遗传因素有关,许多环境条件的变动可影响光合机构的运转,植物的光合机构对它的环境也有一定的适应能力[11]。由于其在植物进化过程中的复杂性而一直成为人们研究植物生理生态的重要内容,因此干旱胁迫下光合作用的变化可以作为判断植物生长和抗逆性强弱的指标,研究干旱胁迫下镜面掌的光合机理,对于镜面掌的在较为缺水的条件下栽培具有一定理论意义和实践意义[12]。本试验研究的主要目的在于探究干旱胁迫条件下镜面掌光合作用指标的变化情况,了解干旱胁迫后复水条件下镜面掌光合生理特性的恢复程度,为镜面掌室内的栽培及养护提供一定的理论基础和科学依据。
1.2植物对干旱胁迫的响应
干旱是影响植物生长和发育的主要环境因子之一,干旱胁迫会使植物产生一系列的生理
- 1 -
干旱胁迫下镜面掌光合作用的动态变化研究
生化变化,如影响到光合作用、物质运输、蛋白质合成和细胞伸长等过程,从而影响到植物生长发育[13]。研究植物的抗旱生理,可以了解植物在干旱胁迫下的生理生化反应及其适应性,为植物抗旱性的鉴定提供理论依据和实践基础。目前,科学家对植物抗旱性研究的方法主要分为两大类:第一,从植物外部形态和内部解剖结构分析总结,研究其与植物抗旱性之间的关系,包括地上和地下部分的生物量、气孔形态等[13]。第二,以植物自身内部的生理生化反应为切入点,测定其相应的生理生化指标,阐明其在植物抗旱性方面的指示和调节作用[13]。这些生理指标主要包括:水分生理指标、光合能力、酶活性及原生质特性、渗透调节、内源激素(如ABA)等[14]。在国内还有许多学者利用稳定碳同位素方法间接测量C3植物的长期水分利用效率,对植物的抗旱性进行比较研究,总结出不同植物的抗旱机理和抗旱性。近年来,随着高新科技和分子生物学领域的发展,发现了部分与植物抵抗旱胁迫相关的物质,如脯氨酸、甘露醇、海藻糖、甜菜碱等,并分离鉴定出编码这些物质的基因以及相关的调控因子,从分子方面揭示了植物抗旱机制。
1.3干旱胁迫对光合作用的影响
光合作用是构成植物生物量的基础,也是植物对水分胁迫最为敏感的生理过程之一[15]。许多学者以净光合速率(Pn)、气孔导度(Gs)、气孔阻力、蒸腾速率(Tr)、胞间CO2浓度(Ci)的变化作为衡量植物光合作用的主要指标,其中净光合速率(Pn)是衡量植物光合作用的即时指标,蒸腾速率(Tr)则是衡量植物蒸腾作用的过程中散失水分的指标,气孔导度(Gs)、气孔阻力和胞间CO2浓度(Ci)是影响或反映C02进入叶肉细胞的重要因素 [15]。
植物的光合作用不但与植物自身的遗传因素密切相关,也受到空气相对湿度和土壤水分含量、光照强度等一系列外界环境因素的影响[16],干旱会使植物的光合能力发生明显的改变,植物光合作用与环境水分因子之间有一定的适应性变化[17]。干旱胁迫造成植物处于水分逆境,不利于植物进行自身光合作用,未能使光能得到最为有效的利用,从而降低植物光合作用的产量[18]。刘霞在研究干旱胁迫对黄栌叶片光合作用的影响中得出结论,在2d~4d干旱的条件下,由于植物自身的调节作用,光合速率下降不明显,而在4d~8d和8d以上干旱条件下,光合作用能力下降较为明显[19]。
根据欧美学者Farquhar和Sharker对水稻光合作用的研究试验后提出的观点,如果净光合速率下降伴随着气孔导度和细胞间隙CO2浓度的下降,则净光合速率下降的主要原因是气孔因素引起的,如果净光合速率下降伴随着气孔导度下降和细胞间隙CO2浓度的上升,则说明净光合速率下降以非气孔因素为主[20],是由于植物细胞本身光合性能下降造成[21]。
土壤的含水量在一定的范围内是影响蒸腾速率的重要因子,干旱胁迫直接影响植物叶片的水分亏缺,通过气孔扩散阻力,影响蒸腾速率。中国科学院植物研究所的学者在研究玉米干旱胁迫下的蒸腾速率变化研究中指出,在干旱胁迫初期,植物的蒸腾速率有所下降,但是下降的不明显,其后随着突然的继续干燥,蒸腾速率几乎呈直线下降,这是由于气孔因子和非气孔因子交替或者综合影响所致[22],。
气孔是植物叶片与外界进行气体交换的主要通道。通过气孔扩散的气体有O2、CO2和水蒸汽。植物在光下进行光合作用,气孔可以根据环境条件的变化来调节自己开度的大小而使
- 2 -
干旱胁迫下镜面掌光合作用的动态变化研究
植物在损失水分较少的条件下获取最多的CO2。气孔开度对蒸腾有着直接的影响,气孔导度和气孔阻力都是描述气孔开度的量,蒸腾作用直接与气孔导度成正比,与气孔阻力呈反比。植物在干旱胁迫下,气孔导度逐渐降低,相反的气孔阻力逐渐增大[23]。
干旱条件下,光合速率的下降与植物对光能和CO2的利用有关系[24]。土壤干旱程度增加,叶片光补偿点、CO2补偿点增高,光饱和点和CO2饱和点下降,这表明干旱胁迫使气孔关闭,叶肉细胞光合活性下降,光能利用率降低[25]。Yordanov等对豆科植物干旱胁迫后复水处理恢复期间的气体交换和叶绿素荧光进行了跟踪测定,结果发现,水分亏缺对豆科植物光合作用的影响非常明显,它导致CO2吸收速率和O2释放速率下降。这一现象在其它绿色植物上也有许多报道[26]。
干旱胁迫对植物光合作用的影响过程是极其复杂的[27]。一般认为干旱胁迫对绿色植物光合作用的影响包括气孔限制和非气孔限制两种因素[28],二者对光合作用的影响因植物种类和干旱胁迫程度的不同而不同,前者是指干旱胁迫使气孔导度下降,CO2进入叶片受阻而使光合下降,从而直接影响植物的光合作用,使植物的生长受阻[29]。随着植物叶片水势的降低,光合速率下降有一个从气孔限制向叶肉细胞光合活性限制的转变过程,也就是向非气孔因素转变的过程,表现为光合器官活性的下降[30]。我国学者许大全在研究小麦叶片光合作用的光转换机制中得出,在干旱胁迫下,引起植物叶片光合效率降低的因素有两类,即气孔限制和非气孔限制,两者有很大的区别,前者使胞间CO2浓度(Ci)降低,而后者使胞间CO2浓度(Ci)增高,当两因素同时存在时,判断哪一个因素占优势,标准应当是胞间CO2浓度(Ci)的变化方向[30]。
干旱胁迫同样影响植物的蒸腾速率。卢苍义等人在研究芦荟等植物在干旱胁迫下的蒸腾特征中指出,在干旱胁迫下,气孔阻力增大,蒸腾速率逐渐降低,水分胁迫越大,蒸腾速率下降的越明显,当下降到临界点时,蒸腾速率将至最低[31]。
- 3 -
干旱胁迫下镜面掌光合作用的动态变化研究
2 试验材料与方法
2.1试验材料
供试验的材料为温室盆栽镜面掌Pilea peperomioides Diels。
2.2试验设计
本项研究在温室盆栽人工控制水分条件下,测定干旱处理不同天数下镜面掌的各项光合指标,目的在于探讨镜面掌在不同干旱天数处理后光合作用指标的变化。
试验采用盆栽法在东北农业大学园艺试验站、园艺学院实验室进行。将扦插于东北农业大学园艺学院智能温室苗盆中,盆的大小为28cm×20cm,有渗水孔,每盆装基质3.5kg,设3次重复,基质由比例为1:3:3的田园土、蛭石和草炭土组成。有机质含量2.15%,全氮含量0.14%,有效氮含量134.5mg/kg,有效磷含量16.5mg/kg,有效钾含量79.3mg/kg,pH值为
6.5,扦插量为每盆三株约同等年龄和生长量的镜面掌,插后立即浇透水正常管理至镜面掌正常生根,更换新叶,生长整齐。环境昼夜温度为25+2℃/18+2℃,相对湿度为65—70%。每天浇透水1次,正常生长后90天进行干旱处理。试验开始时间后第0d充分灌溉,让其自然失水,分别在4d、8d、12d、16d,4次采样用LI—6400测定对应的对照与干旱处理的镜面掌叶片的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)等光合特性指标。然后通过浇水解除干旱胁迫,复水1d后用LI—6400对各项光合指标进行测定。20xx年12月初开始试验,试验材料按照干旱胁迫天数分为5组:
对照:ck(每天浇透水1次);
T1:4d干旱胁迫;
T2:8d干旱胁迫;
T3:12d干旱胁迫;
T4:16d干旱胁迫;
f1: 4d干旱胁迫后复水;
f2: 8d干旱胁迫后复水;
f3: 12d干旱胁迫后复水;
f4: 16d干旱胁迫后复水;
每个处理重复4次。
2.3试验内容与方法
2.3.1光合生理指标动态变化的测定
采用美国LI-COR公司生产的LI-6400便携式光合仪[32],在6400-02B人工光源下,光合有效辐射设定为1400μmol CO2 m-2·s-1,开放气路,空气流速为0.5μmol·s-1,对镜面掌叶片的
- 4 -
干旱胁迫下镜面掌光合作用的动态变化研究
净光合速率(Pn,μmol CO2 m·s)、蒸腾速率(Tr,mmol H2O m·s)、气孔导度(Cond,mol H2O m-2·s-1)、胞间CO2浓度(Ci,μmol CO2 m-2)[31]等指标进行测定。每次每个处理选择4株,测定选择从顶部数起第二节的叶片,每个叶片均连续读取4个数据,最后对所有数据进行处理并利用相关软件对相关指标进行统计分析及对分析结果进行相关的讨论[32]。测定时界环境条件为:温度(TC)25~38℃,大气湿度(RH%)50%,太阳辐射(PAR)500~550μmol·CO2 m-2·s -1。
2.4数据处理与分析
试验结果用Excel软件进行数据处理、分析和绘图。
- 5 -
干旱胁迫下镜面掌光合作用的动态变化研究
3 结果与分析
3.1干旱胁迫条件下镜面掌叶片净光合速率(Pn)的变化
净光合速率(Pn)是评价植物适应性强弱的重要指标,它与植物的生长密切相关[28]。图3-1为干旱胁迫及复水条件下镜面掌叶片净光合速率(Pn)的变化情况。
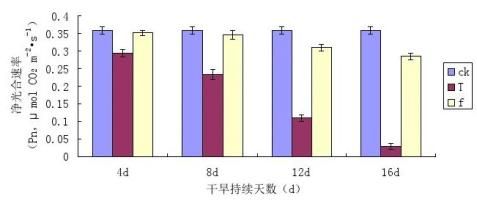
图3-1干旱胁迫及复水条件下镜面掌叶片净光合速率(Pn)的变化 Fig 3-1 The changes of Pilea peperomioides Diels Net photosynthetic rate (Pn) under drought stress
and rehydration
从图3-1中可以看出,随着干旱胁迫天数的增加,镜面掌叶片净光合速率(Pn)的下降幅度逐渐增大,干旱胁迫处理使镜面掌叶片净光合速率Pn均低于对照,T1、T2、T3和T4降低幅度分别是CK的23.08%、45.41%、50.33%、81.18%,T1下降的幅度最小,T4下降幅度最大。说明干旱胁迫程度越大,对镜面掌光合作用的抑制作用越强。胁迫后进行复水处理,复水后各处理镜面掌叶片净光合速率(Pn)与其相对应胁迫处理的净光合速率(Pn)相比均有所回升,TI与f1、T2与f2、T3与f3、T4与f4之间有差异。各处理复水后的增加幅度是其对应的胁迫处理的17.46%、30.77%、40.01%、45.21%,说明光合作用对水分很敏感,胁迫程度越大,复水后的净光合速率恢复的幅度越大。
3.2干旱胁迫条件下镜面掌叶片蒸腾速率(Tr)的变化
干旱胁迫及复水条件下镜面掌叶片蒸腾速率(Tr)的变化情况见图3-2。
- 6 -
干旱胁迫下镜面掌光合作用的动态变化研究
图3-2干旱胁迫及复水条件下镜面掌叶片蒸腾速率(Tr)的变化
Fig 3-2 The changes of Pilea peperomioides Diels transpiration rate (Tr) under drought stress and
rehydration
由图3-2可以看出,不同梯度干旱胁迫处理均使镜面掌叶片蒸腾速率(Tr)均低于对照,随着胁迫程度的加深,蒸腾速率(Tr)均有明显的下降趋势,且胁迫天数越多,蒸腾速率(Tr)越慢,T1、T2、T3和T4的降低量分别是对照的44.99%、52.41%、71.26%、90.53%,说明干旱胁迫对镜面掌叶片蒸腾速率有影响,是导致光合作用降低的另一原因。胁迫后进行复水处理的镜面掌叶片蒸腾速率(Tr)与其相对应的胁迫处理复水前蒸腾速率(Tr)相比均有所回升,TI与f1、T2与f2、T3与f3、T4与f4之间有差异。各处理复水后的增加量分别是其对应的胁迫处理复水前的41.09%、58.41%、60.38%、74.23%,说明干旱胁迫持续天数越多,镜面掌叶片蒸腾速率(Tr)下降的越多,复水后植株的蒸腾速率恢复的越多。
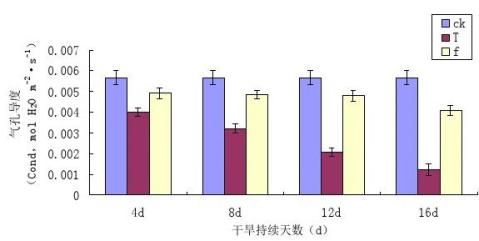
3.3干旱胁迫条件下镜面掌片气孔导度(Gs)的变化
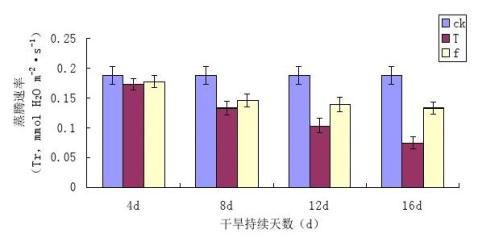
气孔导度(Gs)反映了气孔的开张程度,而气孔的开张程度则直接影响到植物对空气中CO2的利用[33]。图3-3为干旱胁迫及复水条件下镜面掌叶片气孔导度(Gs)的变化情况。
- 7 -
干旱胁迫下镜面掌光合作用的动态变化研究
图3-3干旱胁迫及复水条件下镜面掌叶片气孔导度(Gs)的变化
Fig 3-3 The changes of Pilea peperomioides Diels stomatal conductance (Gs) under drought
stress and rehydration
从图3-3中可以看出,与干旱胁迫对镜面掌叶片净光合速率(Pn)相似,随着胁迫程度的加深,叶片Gs有明显的下降趋势,且胁迫程度越大,Gs越低,不同程度的干旱胁迫使镜面掌叶片气孔导度Gs均低于对照,T1、T2、T3和T4降低幅度分别是对照的70.03%、83.48%、85.02%、92.48%,。胁迫后进行复水处理,各处理镜面掌叶片Gs与其相对应的胁迫处理复水前的Gs相比有所回升,TI与f1、T2与f2、T3与f3、T4与f4之间有差异。各处理复水后的增加幅度是其对应胁迫处理的35.46%、50.33%、70.77%、75.11%。通过分析表明,水分对镜面掌片Gs有着极其敏感的作用,随着干旱胁迫程度的加重,Gs有着剧烈变化。干旱胁迫直接影响到气孔的通透性,影响到H2O和CO2的运输,从而影响植物的光合作用。
3.4干旱胁迫条件下镜面掌叶片气孔阻力(Rs)的变化
干旱胁迫及复水条件下镜面掌叶片气孔阻力(Rs)的变化情况见图3-4。
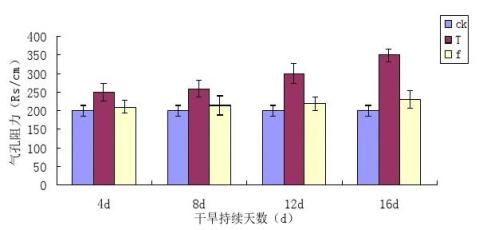
图3-4干旱胁迫及复水条件下镜面掌叶片气孔阻力(Rs)的变化
- 8 -
干旱胁迫下镜面掌光合作用的动态变化研究
Fig 3-4 The changes of Pilea peperomioides Diels stomatal resistance (Rs) under drought
stress and rehydration
通过图3-4可知,干旱胁迫处理下,镜面掌气孔阻力增大,随着胁迫程度的加深,Rs有明显的上升趋势,且胁迫程度越大,Rs越高。不同梯度干旱胁迫处理使镜面掌叶片的气孔阻力Rs均高于对照,T1、T2、T3和T4升高的幅度是对照的30.11%、42.41%、52.40%、83.55%,说明干旱胁迫使气孔的阻力增大,气孔阻力升高是导致光合作用降低的一个原因。胁迫后进行复水处理,各处理镜面掌叶片Rs与其相对应的胁迫处理Rs均有所回升,TI与f1、T2与f2、T3与f3、T4与f4之间有差异。各个处理复水后的增加幅度是其对应的胁迫处理的23.09%、25.01%、26.93%、33.40%,从中可以看出,干旱胁迫的程度越大,复水后镜面掌气孔阻力降低的幅度也越大。
3.5干旱胁迫条件下镜面掌叶片胞间CO2浓度(Ci)的变化
图3-5为干旱胁迫及复水条件下镜面掌叶片胞间CO2浓度(Ci)的变化情况

图3-5干旱胁迫及复水条件下镜面掌叶片胞间CO2浓度(Ci)的变化 Fig 3-5 The changes of Pilea peperomioides Diels intercellular CO2 concentration (Ci) under
drought stress and rehydration
由图3-5可以看出,在不同梯度干旱胁迫处理下,镜面掌叶片Ci均低于对照。在T1处理和T2处理下,Ci表现出随水分含量降低而下降的趋势,而在T3处理下,Ci随干旱胁迫进行不会降低,反而会有所回升,介于T1处理和T2处理之间,Ci表现出随水分含量降低而下降的趋势,随着干旱胁迫程度的加深,Ci下降越明显,T1、T2、T3和T4的降低幅度分别是对照的12.87%、24.83%、17.44%、14.54%,说明气孔开放程度直接降低了叶片内CO2的含量,进而影响叶片的光合作用。胁迫后进行复水处理,各处理镜面掌叶片Ci与其相对应的胁迫处理Ci相比有所回升,TI与f1、T2与f2、T3与f3、T4与f4之间有差异。各处理复水后的增加幅度是其对应的胁迫处理的15.46%、55.77%、40.12%、27.14%。
- 9 -
干旱胁迫下镜面掌光合作用的动态变化研究
4 讨论
4.1干旱胁迫下气孔结构指标与气孔导度(Gs)、蒸腾速率(Tr)及光合速率(Pn)的关系
前人在小麦上研究发现,叶片气孔导度与蒸腾速率、光合速率均呈显著或极显著正相关
[34]。以此说明,器官蒸腾强弱与气孔导度同步变化。本试验结果表明:干旱胁迫条件下,不同干旱程度光合特性有明显差异,轻度干旱处理条件下,镜面掌叶片净光合速率和蒸腾速率下降幅度较小;重度干旱处理条件下,镜面掌叶片的净光合速率和蒸腾速率下降幅度很大,随着干旱胁迫程度的加重,镜面掌叶片净光合速率和蒸腾速率与气孔导度下降趋势越来越明显,说明镜面掌净光合速率和蒸腾速率的下降与气孔导度的下降有直接关系,干旱使镜面掌叶片的气孔部分关闭,促使气孔导度降低,这一方面使通过气孔蒸腾损失的水分减少,另一方面使通过气孔进入叶片的CO2减少,导致蒸腾速率和净光合速率降低,说明气孔限制是干旱胁迫下光合作用下降的主要原因之一[19]。干旱胁迫初期,镜面掌胞间CO2浓度(Ci)的变化趋势与净光合速率(Pn)和气孔导度(Gs)的一致,随胁迫时间的延长而下降;当轻度干旱胁迫8d开始Ci不再下降,而是随胁迫时间的延长逐渐上升而积累。因此,干旱胁迫初期,镜面掌光合下降的原因是气孔导度(Gs)下降导致的气孔限制,当胁迫程度进一步加剧时,气孔限制和非气孔限制共同起作用,逐渐非气孔限制成为主要因子[35]。
4.2干旱胁迫对镜面掌叶片光合特性的影响分析
根据前人的研究结果可知干旱胁迫对绿色植物光合作用的抑制影响包括气孔限制和非气孔限制两类[35],前者是指干旱胁迫使气孔导度下降,CO2进入叶片受阻而使光合作用的能力下降[31],从而直接影响植物的光合作用,使植物的生长受阻。后者是由于干旱使植物叶片叶绿体结构发生变化,植物膜系统遭受损伤,膜脂过氧化加剧而产生超氧自由基,光合色素严重降解,光合电子传递系统遭到破坏,合成酶活性下降、水解酶活性上升,这一系列的生理生化反应致使光合速率下降[36]。本试验结果表明,在干旱胁迫条件下,镜面掌叶片净光合速率与其气孔导度呈显著正相关,都是随土壤含水量的降低而降低。随着土壤含水量的下降,植株与外界水分和气体交换及光合产物的生成速率均受到抑制,并可导致植株水分的失调和光合生理机能的混乱。在干旱胁迫初期,植物根系吸水阻力增强,水分传输能力下降,促使气孔迅速关闭,减少蒸腾失水,以达到植物“节流”的目的,从而导致气体进出气孔阻力增大,蒸腾速率和光合速率都下降,Tr的下降又将导致根系吸水原动力的降低,从而影响根系的水分吸收。此时光合作用下降的主要原因是根系导水能力下降所引起的气孔关闭,即将此时干旱胁迫条件下光合作用下降的原因归结为气孔限制。随着干旱胁迫程度的加深,供试镜
- 10 -
干旱胁迫下镜面掌光合作用的动态变化研究
面掌叶片的Gs、Pn、Tr随之持续降低,Ci逐渐累积。此时叶片光合作用的降低则主要是由于植株水分失调所引起的光合功能紊乱和细胞结构的损伤造成的,即主要是由非气孔限制导致的光合能力的下降。经历干旱胁迫后进行复水1d,各供试镜面掌叶片的Pn、Gs、Tr、Ci开始明显恢复,但各参数均未恢复到CK水平。
在干旱胁迫条件下,镜面掌叶片光合作用受到了显著影响。轻度干旱胁迫时,气孔处在关闭或半关闭状态,光合作用下降的原因主要是由于气孔的开张限制细胞间隙CO2浓度,即气孔限制起主要作用。严重胁迫时,由于植株水分失调所引起的其他生理机能紊乱与反应机构的损伤是其物质代谢受阻的主要原因,亦是决定其光合代谢等生理过程的决定因素。这与前人的研究结论基本一致但与房玉林的研究有所不同[10]。因此可以认为,在轻度或中度干旱胁迫时,气孔限制对植物的光合作用起着主要限制作用,严重干旱胁迫时,则以非气孔限制为主。因此,气孔限制和非气孔限制对光合作用影响的大小取决于干旱胁迫的程度。
- 11 -
干旱胁迫下镜面掌光合作用的动态变化研究
5 结论
干旱胁迫使镜面掌净光合速率(Pn)、蒸腾速率(Tr)和气孔导度(Gs)均有显著降低,气孔阻力(Rs)显著上升,胞间CO2浓度(Ci)先降低后上升的变化,干旱胁迫4d的变化幅度最小,16d的最大。干旱胁迫复水后,各处理的指标都有所回升,且干旱胁迫程度越大,复水后恢复的幅度也越大,但各参数均没有恢复到胁迫前的水平。
在轻度干旱胁迫和中度干旱胁迫处理下,干旱胁迫对镜面掌光合作用的抑制主要是由气孔限制因素引起的,在严重干旱胁迫处理下,镜面掌光合作用的下降原因主要是由于叶片内部结构遭到破坏,即非气孔因素导致的。
综合干旱胁迫及复水条件下的5个光合指标的变化情况得出,干旱胁迫削弱了镜面掌的光合作用,复水后,光合作用能力得到明显恢复,水分对镜面掌光合作用的影响较大。
- 12 -
干旱胁迫下镜面掌光合作用的动态变化研究
参考文献
[1]张其德.干旱胁迫对光合作用的影响[J].北方园艺,2010(18):50-52.
[2]黄颜梅,张健,罗承德.树木杭旱性研究综述[J].四川农业大学学报,1997(1):49-54.
[3]何玉惠.两种驼绒藜属植物抗旱性生理生化指标研究[D].甘肃:甘肃农业大学,2005.
[4]杨成一,刘丛强,宋照亮,等.贵州喀斯特山区植物叶片碳同位素组成研究[J].地球与环境,2007,35(1):33-38.
[5]朱旗,徐吉臣.植物抗旱分子机制研究进展[J].安徽农业科学,2010,38(26):14198-14202,14205.
[6]汤章城.植物对水分胁迫的反应和适应性[I].植物对干旱的反应和适应性[J].植物生理学通讯,1983(4):1-7.
[7]Nobel PS.Adaption of Plant to water and high temperature stress[M].NewYork:John Wiley& sons,1980,43-45.
[8]关义新,戴俊英,陈军,等.土壤干旱下玉米叶片游离脯酸的积累与抗旱性关系[J].玉米科学, 1996,4(1): 43-45.
[9]廖建雄,王根轩.谷子叶片光合速率日变化及水分利用率[J].植物生理学报,1999,25: 362-368.
[10]Kursar T A, Coley P D. Convergence in defensesyndromes of young leaves in tropical rainforests[J]. Biochemical Systematics and Ecology,2003,31:929-949.
[11]吴永美,吕炯章,王书建,李润植.植物抗植物光合作用的机理,2008,28(2):90-93.
[12]焦旭亮,张振文,惠竹梅.土壤逐渐干旱下玉米幼苗蒸腾速率变化的研究.中国科学院植物研究所,2009.
[13]裴冬,张喜英,王竣.高粱、谷子根系发育及其抗旱性研究[J].中国生态农业学报,2002,10(4): 28-30.
[14]孙强等.草地早熟禾草坪土壤水分动态与根系生长分布[J].生态学报,2005,25(6):1307.
[15]梁银丽,陈培元.旱地小麦品种的特征、特性[A]见:山伦、陈培元主编旱地农业生理生态基础.科学出版社,1998,259-264.
[16]韩建秋,王秀峰,张志国.土壤水分梯度对光合作用和根系分布的影响[J].生态学报,2008,28(2):493-499.
[17]HuangBR,FryJD. Root anatomieal,physiological,and morphological reponses to drought stress for tall fescue cultivars [J].Crop Seienee,1998(38).
[18] O’TooleJC,WL Bland.Genotypic variation in crop plan root system[J].Adr.Agron,1997
(41):91-145.
[19]Sharp RE,WJ Davies.Regulation by roots and shoots of water stressd maize Planta[J] Planta,1989(147):43-49.
- 13 -
干旱胁迫下镜面掌光合作用的动态变化研究
[20]Carrow RN.Drought avoidance eharaeteristies of diverse tall fescue cultivars[J]Crop Science,1996(36):371-377.
[21]Deans JD. Fluetuation of the soil environment and fine root growth in a yong sitka sprucePlantation[J].PlantSoil,1979(52):195-208.
[22]HuangBR,Hong WenGao.Root physiological characteristics associated with drought resistance in tall fescue cultivars[J].CropSeienee,2000(40):196-203.
[23]Huang BR, RR Dunean, RN Carrow. Drought resistance mechanism of seven warm season turfgrass under surface soil drying:Ishoot response「J].Crop Seience,1997(38):1558-1863.
[24]Sharp RE,WJ Davies.Root growth and water uptake maize plants in drying soil[J].Exp Bot,1985(36):1441-1456.
[25]Sheffer KM,JH Dunn,DD Minner.Summer drought response and rooting depth of threecool-season turfgrass[J].HortSeience,1987 (22):296-297.
[26]于凤芝,王晓军,邢珊珊,等.4个三叶草种质材料的抗逆性比较研究.草原与草坪,2010,30(2):86-88
[27]LIN Wanhuang,HOFMANN,Rainer,STILWELL Stephen.Physiological responses of five species of trifolium to drought stress.Chin J Appl Environ Biol(应用与环境生物学报)2011,17(4): 580-583.
[28]韩建秋.水分胁迫对光合特性的影响[J].上海应用技术学院学报,2009,9(2):106-109.
[29]殷秀杰,王明玖,石凤翎,崔国文.三种植物幼苗期抗旱性差异的研究. 中国草地学报,2008,30(2):68-72.
[30]马宗仁,刘荣堂.牧草抗旱生理的基本原理[M].兰州:兰州大学出版社,1993.
[31]曾汉来等.植物生理学精品课程.华中农业大学植物科学技术学院,2009.http://nhjy./kech/zwsl/kcjj/index01.asp
[32]何宝坤,李德全.植物渗调蛋白的研究进展[J].生物技术通报,2002(2):6-10.
[33]Regan KL.Evaluation of chemical desiccation as a selection technigne for drought resistance in dryland wheat breeding program[J].Agric Res,1993(44):1683-1691.
[34]李明,王树香,高宝嘉.对不同光照条件下植物生长发育的研究.湖北农业科学,2011,50(14):2921-2923.
[35]李磊,贾志清,朱雅娟,綦艳林.我国干旱区植物抗旱机理研究进展.中国沙漠,2010,30(5):1053-1057.
[36]王丁,张丽琴,薛建辉.林木对干旱胁迫的生理与分子响应研究综述.安徽农业科学, 2011,39(25):15426-15431,15445.
- 14 -
干旱胁迫下镜面掌光合作用的动态变化研究
致谢
本实验在陈雅君老师的悉心指导下完成,从论文的选题、实验设计、结果分析到论文的撰写无不倾注了陈老师的大量心血。四年来,恩师在学习和生活中对我的教导令我终身难忘。恩师不仅在学业上对我严格要求,还注重对我人格的培养,恩师潜移默化传授给我的勇于进取,不怕困难、一丝不苟的精神将会永远伴随着我。
在实验过程中还得到园林实验中心张璐老师的帮助以及园林系其他老师的指导,以及各位师兄师姐的帮助,在此也向他们表示我衷心的感谢。同时也向园林系所有的老师们说一声老师辛苦了,谢谢你们这四年来对我的帮助和支持,这将是我人生旅途中很宝贵的一段经历。
最后,向所有默默关心我的友人和亲人致谢,你们永远是我前进的动力。
- 15 -
-
论文摘要范文 论文摘要的写作方法和范文
论文摘要范文论文摘要的写作方法和范文论文摘要在论文中占据重要的作用是开启全篇论述概述论文内容的关键因此代写毕业论文或论文发表应尤其…
-
论文写作要求及格式范文
论文写作要求及格式范文1论文写作要求11题名准确精炼清晰充分反映论文的基本内涵与特色中英文题名内容应一致英文题名通常以名词短语为主…
-
论文写作要求及范文 (1)
毕业论文设计写作要求一论文格式要求1标题一级标题三号字黑体二级标题小三号字黑体三级标题四号字宋体2目录WORD文档自动生成目录四号…
-
议论文范文与写作运用总汇(1)
议论文范文与写作运用总汇1让纪念闪耀理性光芒纪念是内心情感的涌动但又不是感情的无节制挥霍纪念需要行动来升华但又需要理性的引导真正的…
-
议论文范文与写作运用总汇(2)
议论文范文与写作运用总汇2感情不是保险绳陕西一考生中国是个人情大国除原有的宗族关系之外又重新发展了诸如情同手足亲如兄弟之类的新感情…
-
光合作用知识点总结
光合作用知识总结知识点一叶绿体中的光合色素的种类分布颜色作用知识点二光合色素提取与分离提取原理叶绿体中的色素能够溶解在有机溶剂故常…
-
光合作用知识点总结
第五章细胞的能量供应和利用第四节能量之源光与光合作用一、主要知识点回顾2、色素提取和分离实验注意事项:⑴、丙酮的用途是提取(溶解)…
-
高中生物必修一关于光合作用的总结和解析
高中生物必修一关于光合作用的总结和解析这是高中的重点内容光反应在有光照的条件下植物叶片中叶绿体的类囊体薄膜吸收光能将水分解这叫做水…
-
光合作用知识点总结
54能量之源光与光合作用一光合作用绿色植物通过叶绿体利用光能把二氧化碳和水转化成储存着能量的有机物并释放出氧气的过程叶绿体是进行光…
-
呼吸作用和光合作用总结
第三节ATP的主要来源细胞呼吸一探究酵母菌细胞呼吸的方式1酵母菌一种单细胞真菌在有氧和无氧的条件下都能生存属于兼性厌氧菌2CO2多…