质粒DNA的提取、纯化与鉴定
分子生物学实验报告
题目:质粒DNA的提取、纯化与鉴定
姓名:学号: 班级: 时间:
一、 实验目的:
1. 学习并掌握凝胶电泳进行DNA的分离纯化的实验原理。
2. 学习并掌握凝胶的制备及电泳方法。
3. 学习并掌握凝胶中DNA的分离纯化方法。
4. 掌握碱变性提取发的原理及各种试剂的作用。
5. 掌握碱变性法提取质粒DNA的方法。
二、 实验原理:
1. 质粒DNA的提取——碱变性提取法:
提取和纯化质粒DNA的方法很多,目前常用的有:碱变性提取法、煮沸法、羟基磷灰石柱层析法、EB-氯化铯密度梯度离心法和Wizard法等。其中,碱变性提取法最为经典和常用,适于不同量质粒DNA的提取。该方法操作简单,易于操作,一般实验室均可进行。提取质粒DNA纯度高,可直接用于酶切、序列测定及分析。EB-氯化铯密度梯度离心法,主要适合于相对分子质量与染色体DNA相近的质粒,具有纯度高、步骤少、方法稳定,且得到的质粒DNA多为超螺旋构型等优点,但提取成本高,需要超速离心设备。少量提取质粒DNA还可用沸水浴法、Wizard法等,沸水浴法提取的质粒DNA中常含有RNA,但不影响限制性核酸内切酶的消化、亚克隆及连接反应等。 碱变性法提取质粒DNA一般包括三个基本步骤:培养细菌细胞以扩增质粒;收集和裂解细胞;分离和纯化质粒DNA。
在细菌细胞中,染色体DNA以双螺旋结构存在,质粒DNA以共价闭合环状形式存在。细胞破碎后,染色体DNA和质粒DNA均被释放出来,但两者变性与复性所依赖的溶液pH值不同。在pH值高达12.0的碱性溶液中,染色体DNA氢键断裂,双螺旋结构解开而变性;共价闭合环状质粒DNA的大部分氢键断裂,但两条互补链不完全分离。当用pH值4.6的KAc(或NaAc)高盐溶液调节碱性溶液至中性时,变性的质粒DNA可恢复原来的共价闭合环状超螺旋结构而溶解于溶液中;但染色体DNA不能复性,而是与不稳定的大分子RNA、蛋白质-SDS复合物等一起形成缠连的、可见的白色絮状沉淀。这种沉淀通过离心,与复性的溶于溶液的质粒DNA分离。溶于上清的质粒DNA,可用无水乙醇和盐溶液,减少DNA分子之间的同性电荷相斥力,使之凝聚而形成沉淀。由于DNA与RNA性质类似,乙醇沉淀DNA的同时,也伴随着RNA沉淀,可利用RNase A将RNA降解。质粒DNA溶液中的RNase A以及一些可溶性蛋白,可通过酚/氯仿抽提除去,最后获得纯度较高的质粒DNA。
2. 凝胶电泳进行DNA分离纯化:
电泳(electrophoresis)是带电物质在电场中向着与其电荷相反的电极方向移动的现象。各种生物大分子在一定pH条件下,可以解离成带电荷的离子,在电场中会向相反的电极移动。凝胶是支持电泳介质,它具有分子筛效应。含有电解液的凝胶在电场中,其中的电离子会发生移动,移动的速度可因电离子的大小形态及电荷量的不同而有差异。利用移动速度差异,就可
1
以区别各种大小不同的分子。因而,凝胶电泳可用于分离、鉴定和纯化DNA片段,是分子生物学的核心技术之一。
凝胶电泳技术操作简单而迅速,分辨率高,分辨范围广。此外,凝胶中DNA的位置可以用低浓度荧光插入染料如溴化乙锭(ethidium bromide,EB)或SYBR Gold染色直接观察到,甚至含量少至20pg的双链DNA在紫外激发下也能直接检测到。需要的话,这些分离的DNA条带可以从凝胶中回收,用于各种各样目的的实验。
分子生物学中,常用的两种凝胶为琼脂糖(agarose)和聚丙烯酰胺凝胶。这两种凝胶能灌制成各种形状、大小和孔径,也能以许多不同的构型和方位进行电泳。聚丙烯酰胺凝胶分辨率高,使用于较小分子核酸(5—500bp)的分离和蛋白质电泳。它的分辨率非常高,长度上相差1bp或质量上相差0.1%的DNA都可以彼此分离,这也是采用聚丙烯酰胺凝胶电泳进行DNA序列分析的分子基础。虽然它能很快地进行电泳,并能容纳较大的DNA上样量,但是与琼脂糖凝胶相比,在制备和操作上繁琐。琼脂糖是从海藻中提取的长链状多聚物,由β-D-吡喃半乳糖与3,6-脱水-L-吡喃半乳糖组成,相对分子质量为104-105。琼脂糖加热至90℃左右,即可溶化形成清亮、透明的液体,浇在模版上冷却后形成凝胶,其凝固点为40-45℃。琼脂糖凝胶相对于聚丙烯酰胺凝胶分辨率低,但它的分离范围更大(50至百万bp),小片段DNA(50-20000bp)最适合在恒定轻度和方向的电场中水平方向的琼脂糖凝胶内电泳分离。琼脂糖凝胶电泳易于操作,适用于核酸电泳,测定DNA的相对分子质量,分离经限制酶水解的DNA片段,进一步纯化DNA等。
琼脂糖凝胶电泳是一种常用的方法。在溶液中,由于核酸有磷酸基而带有负电荷,在电场中向正极移动。DNA在琼脂糖凝胶中的电泳迁移率主要取决于6个因素:样品DNA分子的大小、DNA分子的构象、琼脂糖浓度、电泳所用电场、缓冲液和温度。
三、 主要仪器和材料试剂:
1. 仪器和材料:
恒温振荡培养箱,高速冷冻离心机,旋涡振荡器,水浴锅,1.5mL离心管,50mL离心管,不同型号的吸头,微量移液器,微波炉,电泳仪,制胶槽,电泳槽,梳子,锥形瓶,电子天平,手套,紫外灯,Eppendorf管等。 菌体:E.coli DH5α受体菌,具有Ampr标记的质粒pUC19。
2. 实验试剂:
LB培养基,抗生素(氨苄青霉素),溶液Ⅰ,溶液Ⅱ,溶液Ⅲ,RNase A母液,TE缓冲液,饱和酚,氯仿/异戊醇混合液,酚/氯仿/异戊醇(PCI)混合液,预冷无水乙醇,TAE电泳缓冲液(10×),上样缓冲液(6×),琼脂糖,溴化乙锭(EB),DNA相对分子质量标准物DNA Marker λ/Hind Ⅲ,5mol/L pH 5.2的醋酸钠。
四、 实验步骤:
1. 菌体培养:
(1) 配制40mL液体LB培养基(加入5%葡萄糖)、100mL固体LB
培养基、并准备足量的移液管、200μl微量移液器头、1000μl微量移液器头、50mL离心管、1.5mL离心管,灭菌备用。
(2) 向液体LB培养基移取32μl氨苄青霉素,混合均匀。
(3) 将提前活化的E.coli DH5α受体菌,接种于液体LB培养基中。
2
将锥形瓶放入恒温震荡培养箱中,37℃,200r/min培养12-16h。
2. 质粒DNA的提取:
(1) 称量50mL离心管重量W1,取30mL菌液于已称重的50mL离心
管中,配平后6000r/min离心5min。
(2) 弃上清,向离心管中加入5mL溶液Ⅰ,涡旋振荡,6000r/min
离心5min。
(3) 弃上清并称重W2,求W=W2-W1。
(4) 按照1.0mL/100mg菌体的量加入溶液Ⅰ,充分涡旋振荡,冰浴
5min,再按照2.0mL/100mg菌体的量加入溶液Ⅱ,温和颠倒混匀,冰浴2min,然后再按照1.5mL/100mg菌体的量加入溶液Ⅲ,温和颠倒混匀,冰浴10min,平衡后12000r/min离心15min。
(5) 取上清至新50mL离心管内,记录体积,加入两倍体积的冰乙
醇,混匀后在-20℃环境下保存30min。取出后12000r/min离心15min,弃上清,加入5mL70%乙醇,12000r/min离心5min,弃上清,加入5mL70%乙醇,12000r/min离心5min,弃上清,37℃放置5-10min。
(6) 取出离心管,加入1mLTE溶液得到粗提物,加入RNase A液,
使溶液浓度为150μg/mL,37℃保存60-120min。
3. 质粒DNA的纯化:
(1) 取500μl粗提物于1.5mL离心管中,加入等体积的Tris饱和酚,
混匀,12000r/min离心10min。
(2) 转移上清(体积V1)至新管,加入等V1的酚:氯仿:异戊醇溶液,
混匀,12000r/min离心5min。
(3) 转移上清(体积V2)至新管,加入等V2的氯仿:异戊醇溶液,混
匀,12000r/min离心5min。
(4) 转移上清(体积V3)至新管,加入1V3的3M NaAc溶液(pH5.2),10
再加入2V4(V4=V3+13)的冰乙醇,混匀,-20℃保存30-60min,10
取出后12000r/min离心15min。
(5) 弃上清,加入500μl 70%乙醇,10000r/min离心2min,再加入
500μl 70%乙醇,10000r/min离心2min,37℃保存5-10min。
(6) 取出离心管,向一只离心管中加入25μlTE溶液,溶解沉淀后,
转移入另一只离心管中,再取25μl TE溶液加入第一只离心管中,溶解后再移入另一只离心管中,得到50μl纯化质粒DNA。
4. DNA纯度检测:
(1) 取40mLTAE(1×)于300mL锥形瓶中,加入0.4g琼脂糖,放入微
波炉内使其熔化,60℃时倒入准备好的制胶槽中。
(2) 取5.0μl纯化DNA加入1.0μl上样缓冲液,混合,进行点样。
(3) 点样完毕后,100V,200mA条件下电泳30min。
(4) 电泳完毕后,进行EB染色,用凝胶成像仪拍照,得到实验结
果。
3
五、 实验结果:
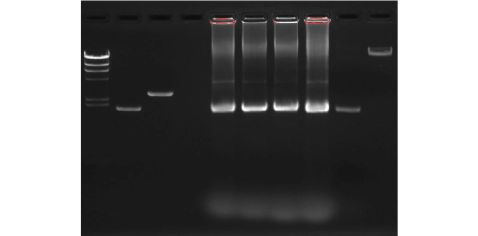
凝胶成像仪拍照如下:
图1 DNA纯度检测凝胶成像结果(1号泳道:DNA markerλ/HindⅢ;2号泳道:超螺旋状态的pUC19质粒DNA;3号泳道:线性的pUC19质粒DNA;4号泳道:空泳道;5至9泳道:7至11组pUC19质粒DNA样品;10号泳道:λDNA)
本组点样在第9泳道,照片显示各种杂质去除得较为彻底,得到了较高纯度的超螺旋状态的pUC19质粒DNA。
六、 讨论:
1. 为获得高纯度的质粒DNA,必须彻底去除杂蛋白、染色体DNA和RNA。在整个质粒提取过程中出去染色体DNA的关键步骤是加入溶液Ⅱ、溶液Ⅲ的变性和复性环节,应控制好变性和复性的时机。加入溶液Ⅰ时,可剧烈震荡,使菌体沉淀转变成均匀的菌悬液,此时细胞尚未破裂,染色体不会断裂;加入溶液Ⅱ时,菌液变粘稠、透明,无菌块残留;加入溶液Ⅲ时,会立即出现白色沉淀。加入溶液Ⅱ和溶液Ⅲ后,应缓慢上下颠倒离心管数次,切忌在旋涡振荡器上剧烈振荡,否则染色体DNA会断裂成小片段,不形成沉淀,而溶解在溶液中,与质粒DNA混合在一起,不利于质粒DNA提纯。因此,操作时一定要缓慢柔和,采用上下颠倒的方法,既要使试剂与染色体DNA充分作用,又不破坏染色体的结构。
2. 酚具有腐蚀性,能损伤皮肤和衣物,使用时应小心。皮肤如不小心沾到酚,应立即用碱性溶液、肥皂或大量清水冲洗。
3. 为最大限度去除上清,可在倒掉部分上清后,再将离心管放入离心机稍作离心,使残留在管壁的液体集中到离心管底部,再用移液器移除液体。
4. 注意上样时要小心操作,避免损坏凝胶或将样品槽底部的凝胶刺穿。也不要快速按出吸头内的样品,避免挤出的空气将样品冲出样品孔。
4
5. 本次实验中,原应灭菌之前加入培养基内的葡萄糖由于操作疏忽而忘记加入,随后按照老师指示配制100mL 10%的葡萄糖溶液一起灭菌,待灭菌后再将葡萄糖溶液加入培养基中。
6. 培养菌体所用的14个锥形瓶有2个锥形瓶中没有菌体生长迹象,另外有2个锥形瓶内菌体生长较少。原因可能是接种时操作失误,例如可能是接种环挑取菌种后离酒精灯火焰太近或者挑取菌种前没有充分冷却,也有可能是接种时接入的菌量过少等。
7. 在纯化DNA加入冰乙醇的步骤中,2只离心管在加入的DNA样品量相同的情况下,出现了从-20℃环境下取出后2只离心管内液体体积相差较多的情况,分析后发现量较少的1只离心管内加入的各种试剂的量是合适的。可能的原因:①在用移液器移取冰乙醇时下按力度太大,吸入过多液体;②2只离心管分别由2名组员加液,操作上可能出现个人之间的误差。
8. 本次实验得到的质粒DNA较少,最主要原因是选择了菌体生长较少的培养基,仅得到较少的粗提物,导致最终纯化得到的质粒DNA也较少。
七、 附注:
培养基及实验试剂配方:
1. LB(Luria Broth)培养基
LB液体培养基:1%蛋白胨(typtone),0.5%酵母粉(yeast extract),1%NaCl。用NaOH调pH至7.2, 121℃灭菌20min备用。
LB固体培养基:除以上LB培养基成分外,还需要添加1.5%~2%琼脂,121℃灭菌20min备用。
LB半固体培养基:在LB液体培养基中加入0.75%琼脂,121℃灭菌20min备用。
2. 氨苄青霉素:母液浓度为100μg/μl,工作浓度为50~100μg/mL。
3. TE缓冲液:10mmol/L Tris-HCl(pH 8.0),1mmol/L EDTA。
4. 氯仿/异戊醇混合液:按氯仿:异戊醇为24:1(V/V)的比例在氯仿中加入异戊醇。
5. 酚/氯仿/异戊醇(PCI)混合液:
饱和酚:分析纯的酚溶化后经160℃重蒸后,加入0.1%(V/V)的抗氧化8-羟基喹啉,再加入等体积的0.1mol/L Tris-HCl(pH 8.0)缓冲液反复抽提,使之饱和,并使其pH为8.0,亦可购买商品化的饱和酚。
按酚与氯仿/异戊醇为1:1的比例混合饱和酚与氯仿/异戊醇混合液,即得酚/氯仿/异戊醇(PCI)(25:24:1)混合液。
6. 6×上样缓冲液:30%甘油,0.25%溴酚蓝(bromophenol blue, BPB),0.25%二甲苯氰FF。
7. 溶液Ⅰ:50mmol/L葡萄糖,25mmol/L Tris-HCl(pH8.0),10mmol/L EDTA。
8. 溶液Ⅱ:0.2mol/L NaOH,1%SDS(临用前用10mol/L NaOH和20%SDS母液配制)。
9. 溶液Ⅲ:5mol/L KAc 60mL,冰醋酸11.5mL,重蒸水28.5mL;溶液终浓度为:K+3mol/L,Ac-5mol/L;
10.RNase A 母液:将RNase A溶于含10mmol/L Tris-HCl(pH7.5)和15mmol/L NaCl的溶液中,配成10mg/mL溶液,于100℃加热15min,使污染的DNase失活。冷却后用1.5mL无菌离心管分装,保存于-20℃。
5
第二篇:大肠杆菌质粒DNA的提取
一、大肠杆菌质粒DNA的提取
质粒DNA的提取是从事基因工程工作中的一项基本实验技术,但提取方法有很多种,以下介绍一种最常用的方法:
碱裂解法:此方法适用于小量质粒DNA的提取,提取的质粒DNA可直接用于酶切、PCR扩增、银染序列分析。方法如下:
1、接1%含质粒的大肠杆菌细胞于2ml LB培养基。
2、37℃振荡培养过夜。
3、取1.5ml菌体于Ep管,以4000rpm离心3min,弃上清液。
4、加0.lml溶液I(1%葡萄糖,50mM/L EDTA pH8.0,25mM/L Tris-HCl pH8.0)充分混合。
5、加入0.2ml溶液 II(0.2 mM/L NaOH,1% SDS),轻轻翻转混匀,置于冰浴5 min 。
6、加入0.15m1预冷溶液III(5 mol/L KAc,pH4.8),轻轻翻转混匀,置于冰浴5 min 。
7、以10,000rpm离心20min,取上清液于另一新Ep管
8、加入等体积的异戊醇,混匀后于?0℃静置10min。
9、再以10,000rpm离心20min,弃上清。
10、用70%乙醇0.5ml洗涤一次,抽干所有液体。
11、待沉淀干燥后,溶于0.05mlTE缓冲液中
煮沸法
1、将1.5ml培养液倒入eppendorf管中,4℃下12000g离心30秒。
2、弃上清,将管倒置于卫生纸上几分钟,使液体流尽。
3、将菌体沉淀悬浮于120ml STET溶液中, 涡旋混匀。
4、加入10ml新配制的溶菌酶溶液(10mg/ml), 涡旋振荡3秒钟。
5、将eppendorf管放入沸水浴中,50秒后立即取出。
6、用微量离心机4℃下12000g离心10分钟。
7、用无菌牙签从eppendorf管中去除细菌碎片。
8、取20ml进行电泳检查。
[注意] 1. 对大肠杆菌可从固体培养基上挑取单个菌落直接进行煮沸法提取质粒DNA。
2. 煮沸法中添加溶菌酶有一定限度,浓度高时,细菌裂解效果反而不好。有时不同溶菌酶也能溶菌。
3. 提取的质粒DNA中会含有RNA,但RNA并不干扰进一步实验,如限制性内切酶消化,亚克隆及连接反应等。
质粒DNA的大量提取和纯化
在制作酶谱、测定序列、制备探针等实验中需要高纯度、高浓度的质粒DNA,为此需要大量提取质粒DNA。大量提取的质粒DNA一般需进一步纯化,常用柱层析法和氯化绝梯度离心法。
(一)、碱法
1、取培养至对数生长后期的含pBS质粒的细菌培养液250ml,4℃下5000g离心15分钟,弃上清,将离心管倒置使上清液全部流尽。
2、将细菌沉淀重新悬浮于50ml用冰预冷的STE中(此步可省略)。
3、同步骤1方法离心以收集细菌细胞。
4、将细菌沉淀物重新悬浮于5ml溶液I中,充分悬浮菌体细胞。
5、加入12ml新配制的溶液II, 盖紧瓶盖,缓缓地颠倒离心管数次,以充分混匀内容物,冰浴10分钟。
6、加9ml用冰预冷的溶液III, 摇动离心管数次以混匀内容物,冰上放置15分钟,此时应形成白色絮状沉淀。
7、4℃下5000g离心15分钟。
8、取上清液,加入50ml RNA酶A(10mg/ml), 37℃水浴20分钟。
9、加入等体积的饱和酚/氯仿,振荡混匀,4℃下12000g离心10分钟。
10、取上层水相, 加入等体积氯仿,振荡混匀,4℃下12000g离心10分钟。
11、取上层水相,加入1/5体积的4mol/L NaCl和10% PEG(分子量6000), 冰上放置60分钟。
12、4℃下12000g离心15分钟, 沉淀用数ml 70%冰冷乙醇洗涤,4℃下12000g离心5分钟。
13、真空抽干沉淀,溶于500ml TE或水中。
[注意] 1. 提取过程中应尽量保持低温。
2. 加入溶液II和溶液III后操作应混和,切忌剧烈振荡。
3. 由于RNA酶A中常存在有DNA酶,利用RNA酶耐热的特性,使用时应先对该酶液进行热处理(80℃ 1小时),使DNA酶失活。
二、质粒DNA琼脂糖凝胶电泳鉴定
琼脂糖是从海藻中提取出来的一种线状高聚物,应选用电泳纯的,琼脂糖此级产品筛除了抑制物和核酸酶,而且用溴化乙锭染色后荧光背景最小。
(1)琼脂糖凝胶电泳装置
由于琼脂糖凝胶电泳既要求不高,而适应性又强,在过去15年里已成功地设计了形形色色及大大小小的电泳槽。对这些装置的选择主要是依据个人的喜恶。使用最普遍的装置是Walter Schaffner发明的水平板凝胶。
水平板凝胶通常在一块可安放于电泳槽平台的玻璃板或塑料盘上灌制。在有些装置中,则可将凝胶直接铺在平台上。凝胶恰好浸在缓冲液液面下进行电泳。凝胶的电阻几乎与缓冲液的电阻相同,所以有相当一部分的电流将通过凝胶的全长。
(2)琼脂糖凝胶的制备
琼脂糖凝胶的制备是将琼脂糖在所需缓冲液中熔化成清澈、透明的溶液。然后将熔化液倒入胶模中,令其固化。凝固后,琼脂糖形成一种固体基质,其密度取决于琼脂糖的浓度。通贯凝胶的电场接通后,在中性pH值下带负电荷的DNA向阳极迁移。
(3)琼脂糖凝胶的染色
电泳完毕,将琼脂糖凝胶转移入含EB的染液中,染色10分钟,取出紫外灯下观察。
三、大肠秆菌感受态细胞的制备
感受态的细胞可以摄入外部溶液中的DNA,而常态的细胞却不能,所以要转化质粒DNA进入大肠杆菌必须首先制备感受态的大肠杆菌细胞。
1、取1%大肠杆菌E.coli接种于含2ml LB培养基的试管中,37℃振荡培养过夜
2、取0.1ml过夜培养物转种于含10ml LB培养基的三角瓶中,37℃振荡培养3h至OD600=0.3
3、然后把培养物倒入1.5ml离心管中,冰浴10min。
4、在4℃下以4000rpm离心5min,去上清液
5、把菌体悬浮于15m1冰冷的0.1M CaCl2溶液中,置冰上30min
6、然后再在4℃下以4000rpm离心10min,去上清液
7、将菌体悬浮于0.1ml CaCl2溶液中,冰浴放置4-12hr备用。
四、质粒DNA高频转化大肠杆菌
制备好感受态细胞后,接下来就是质粒DNA转化入大肠杆菌细胞的过程。但要注意的是感受态细胞最好是新制备的,因为保存一定时间的感受态细胞会使转化率降低;此外DNA的浓度也要注意,不能太高。
1、取新制备的一管感受态细胞。
2、取0.03ml感受态细胞转和4ng质粒DNA混匀,置冰浴30min
3、将 Ep管置于42℃水浴中热冲击2分钟,立即置于冰上1分钟。
4、在 Ep管中加70u1 LB培养基混匀,37℃培养30min
5、涂在含适当浓度抗生素的LB平板上。
6、37℃培养过夜,长出的菌斑既为阳性克隆。
五、线形质粒DNA 5'-粘性末端去磷酸
经限制性内切酶酶切的质粒DNA 5'端均带有磷酸集团,如用此载体直接转化大肠杆菌,其自身环化几率非常高,影响连接、转化效率。因此一般酶切的质粒DNA要经脱磷,使用碱性磷酸酶(CIAP)去除5'端磷酸集团。方法如下:
1、质粒DNA和CIAP以及BUFFER 37℃水浴30min
2、补充CIAP 再37℃水浴30min
3、加入50mM EDTA至终浓度5mM 75℃加热10min 失活CIAP
4、冷却至室温,酚-氯仿抽提,乙醇沉淀浓缩。
5、抽干溶液,加适量TE溶解。
六、聚合酶链式反应(PCR)技术
PCR是一种选择性体外扩增DNA或RNA片段的方法。其特异性是由两个人工合成的引物序列决定的。所谓引物就是与待扩增DNA片段两翼互补的寡聚核苷酸,其本质是ssDNA片段。待扩增DNA模版加热变性后,两引物分别与两条DNA的两翼序列特异复性。此时,两引物的3'端相对,5'向背。在合适的条件下,由Taq DNA聚合酶催化引物引导的DNA合成,既引物的延伸。上述过程是由温度控制。这种热变性-复性-延伸的过程就是一个PCR循环。PCR就是在合适条件下的这种循环的不断重复。理论上扩增产物量成指数上升,既n个循环后,产量为2n拷贝。典型的PCR操作过程如下:
1、 反应体系:
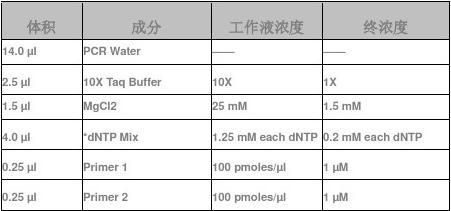

(反应溶液可置于毛细管或薄壁离心管中扩增)
2、操作过程:(所用仪器为AMP1605热循环PCR扩增仪 预变性:94℃ 90秒
循环过程:94℃ 1秒 48℃ 1秒 72℃ 40秒 延伸:72℃ 5分钟
3、检测:琼脂糖电泳检测。 40个循环
-
质粒DNA的提取及其琼脂糖凝胶电泳实验报告及思考题
1实验目的1通过本次实验学习和掌握碱裂解法提取质粒2通过本次实验学习琼脂糖凝胶电泳检测DNA的方法和技术2实验材料及用品1实验仪器…
-
质粒DNA抽提实验报告
质粒DNA抽提实验报告一实验目的1掌握碱裂解法小量快速提取质粒DNA的方法提取的质粒DNA可直接用于酶切PCR扩增等2学习利用水平…
-
实验一、质粒DNA的提取及检测实验报告
实验一质粒DNA的提取及检测实验目的1掌握碱裂解法提取质粒的原理和步骤2掌握琼脂糖凝胶电泳检测DNA的方法和技术3学会PCR操作的…
-
质粒DNA的提取实验报告
质粒DNA的提取一实验方法碱裂解法抽提质粒DNA二实验原理基于质粒DNA与染色体DNA变性与复性的差异三实验步骤1质粒提取1100…
-
质粒的提取酶切 实验报告
实验一质粒的提取酶切实验目的掌握质粒小量快速提取法用琼脂糖凝胶电泳法鉴定其纯度实验原理质粒是一种染色体外的稳定遗传因子大小在120…
-
班主任安全工作总结
五化中学20xx—20xx学年度下学期七年五班安全工作总结孩子的健康、快乐的成长是我们每位老师的职责,在教给他们科学文化知识的同时…
-
在七年级期中考试总结会上的讲话
各位老师,同学们:下午好!,经过我们全体师生的共同努力,本学期的期中考试已圆满结束,在这次考试中一大批同学取得了优异的成绩,如:以…
-
职工培训总结
八连冬季职工培训总结八连二〇〇九年十二月八连冬季职工培训总结今冬,我连党支部根据师团党委统一安排部署,按照团党委关于开展冬春季职工…
-
野外生存协会活动总结
野外生存协会20xx年度下半年总结化学与生命科学学院20xx.12为了全面贯彻党的教育方针,促进学生的健康发展,更好地加强校园的精…
-
演讲比赛活动总结
大学生社团联合会(演讲比赛)活动总结部门:***姓名:***二〇一二年五月活动总结上周,由我们大学生社团联合会举办了演讲比赛,由于…