细胞生物学实验报告
染色体标本的制备及观察
泮力菁 201100140091 20##级生物基地 同组者:商倩倩
【实验目的】
1、掌握染色体标本制作的方法;
2、认识不同生物染色体的特征,学会制作染色体组形图;
【实验原理】
1、制作染色体标本的意义:A、生物学方面:根据染色体特征鉴别生物及种类(例如猫38;小鼠40;大鼠42;兔44;人46;马64;鸡和狗78);B、临床医学方面:主要用于遗传性疾病的诊断及研究(例如21三体综合症,卵巢退化症;睾丸退化症等)。因此,染色体阻性实验广泛应用与胎儿遗传性疾病早期诊断中(抽羊水做组型)。
2、染色体标本的制作原理:A、制作染色体标本的先决条件:细胞具有旺盛的分裂能力(选择活跃的组织:胸腺,骨髓,睾丸,小肠;施加药物使细胞分裂);设法得到大量的分裂中期细胞(秋水仙素)。B、重要点:PHA:促细胞分裂,使淋巴细胞返幼,变为淋巴母细胞;秋水仙素:破坏微管装配,使纺锤体不能形成,使大量细胞停止在分裂中期;低渗作用:水进入细胞内,细胞内容空间变大,染色体间的距离拉大,抑郁cs分散开;空气干燥:使细胞和cs展开;固定:用carnoy’s solution(甲醇:冰醋酸=3:1)作用是蛋白质变性,对染色体内的组蛋白讲,变性后硬度增加,保持了染色体的“即时形态”,对细胞膜蛋白讲,变性使细胞膜硬度增加,形成屏障作用,防止了细胞内物质的外溢和丢失。
3、本实验选择的实验材料是小鼠的睾丸。睾丸是雄性动物生殖细胞发育和成熟的部位,是产生精子的器官。睾丸内的曲细精管从外到内依次分布有精原细胞、初级精母细胞、次级精母细胞、精子细胞以及形态渐变中的精子。取睾丸将曲细精管中的细胞冲出后固定,然后制片染色,可以观察到动物生殖细胞减数分裂染色体的不同时相。如果提前给动物的腹腔注射秋水仙素溶液可抑制生殖细胞的正常分裂,增加了中期分裂细胞的数目,此方法可用来进行减数分裂过程中染色体的计数与观察。
睾丸染色体标本的制备,可以用来检测雄性个体生殖细胞是否受到周围环境污染或是否发生病变,是小型动物生殖学常规实验。
4、染色体标本的制作:
① 观察:由于分裂期中期的染色体染色体凝集程度最高,因而我们应尽量使细胞停留在分裂期中期以便观察。
② 细胞:为尽量看到多的染色体,我们应当选取具有旺盛分裂能力的细胞,这样的细胞可以来自:胸腺、骨髓、睾丸、小肠等。我们在此实验中使用的是小鼠的精巢细胞。在做人的染色体标本时,我们使用的是血液里的淋巴细胞。
③ 药物: PHA(植物细胞凝集素):PHA具有促进细胞凝集和分裂的作用。PHA可以使血液中的淋巴细胞还原至淋巴母细胞(只存在于骨髓中); 秋水仙素:秋水仙素可以破坏微管的组装,纺锤体不能形成,细胞分裂停在中期。与秋水仙素作用相同的药物还有长春花碱、鬼臼素、苯环己烯等。可以促进胃管组装的药物有紫杉酚、重水等。
④ 空气干燥:使细胞与染色体易于分离
⑤ 低渗作用:水进入细胞内,细胞内容空间变大,染色体距离变大,易于分离、
展平。
⑥ 固定:Camoy’s solution,使组蛋白变性(变硬),维持染色体原有形态
5、 描述染色体的四个参数
① 相对长度=(每条染色体的长度)/(单倍常染色体之和+X染色体)*100
相对长度可以用来表示每条染色体的长度
② 臂指数=(长臂的长度)/(短臂的长度) 臂指数可以用来确定臂的长度 ;为了更准确的区别亚中部和亚端部着丝粒染色体1964年Levan提出划分标准:臂指数在1.0-1.7之间,为中部着丝粒染色体(M);1.7-3.0之间,为亚中部着丝粒染色体(SM);3.0-7.0之间,为亚端部着丝粒染色体(ST);7.0以上,为端部着丝粒染色体(T)。
③ 着丝粒指数=(断臂长度)/(染色体全长)*100 ;着丝粒指数可以决定着丝粒的相对位置 ;按Levan的划分标准,着丝粒指数在50.0-37.5之间,为M;37.5-25.0之间,为SM;25.0-12.5之间,为ST;12.5-0.0之间,为T 。
④ 染色体臂数(NF),根据着丝粒的位置来确定 端着丝粒染色体(T),NF=1
中部,亚中部,亚端部着丝粒染色体(M,SM,ST),NF=2
【实验器材】
1、实验材料:
A、雄性小鼠(6-8周龄,20-30g);
B、0.02%秋水仙素(4℃保存);生理盐水;0.3%氯化钾溶液;甲醇·冰醋酸固定液(3:1);Giemsa染液;
2、实验仪器:
小烧杯;剪刀;铜网;离心管;离心机;吸管;低温预冷载玻片;滤纸;普通光学显微镜;恒温培养箱(恒温水浴锅);镊子;注射器;玻璃片;
【实验步骤】
1、取雄性小鼠以每克体重4μg注射秋水仙素,经14~16小时后,断头法杀死小鼠,取出睾丸用生理盐水(0.9%的NaCl)洗去血污。
2、放入装有1ml 0.3%KCl液的小烧杯中剪碎至呈乳白色。
3、用铜网过滤到刻度离心管中,再加0.3%KCl液至4ml。
4、37℃静置30分钟,进行低渗处理。
5、以800~1000转/分离心8分钟。
6、弃上清液,加入2ml甲醇·冰醋酸固定液(3:1),并用吸管至管底吹气,轻轻打散细胞,固定8分钟。
7、再以800~1000转/分离心8分钟。
8、弃上清液,加1ml固定液,再制成细胞悬液,固定5分钟。
9、取洁净的低温预冷载片,距载片10~15cm高度滴下2~3滴细胞悬液,从载片一边向另一边轻轻吹气,并同时轻轻敲打载片,以使细胞均匀分布和促使染色体展开。
10、用滤纸擦去载片上多余液体,空气干燥或成文火干燥。
11、用 Giemsa染色20 ~ 30分钟,细水冲洗玻片背面,去多余染液,气干。
镜检:低倍镜下寻找分散良好、染色适中的分裂相,高倍镜或油镜下观察染色体形态并计数。
【注】:倒置染色法——在玻璃板上用废旧的载玻片做支架,使标本载玻片的标本面向下放置到支架上,在玻璃板和标本载玻片之间滴加Giemsa染液,其目的是节省染料,避免染液快速挥发,放置染色颗粒沉淀,影响观察。在操作时应注意,多个样品同时染色应摆放紧密,不要有间隙;地然也是应尽量慢,不要有气泡,以免部分染色体不被着色。
【实验结果】
1、实验图片


图1:小鼠精细胞 (10*40) 图2:中期染色体(10*40)
【分析】由图中可以看出,小鼠睾丸内的精细胞较中期染色体大,而且一端较尖,并没有染色体的形态;图2中的染色体约为64条,大于小鼠一个细胞中的染色体数(40条)。


图3:中期染色体(放大) 图4中期染色体(放大)
【分析】图3中的染色体分离较好,但是背景较杂;图4中背景干净清晰,但是染色体形态分析并没有完全进入中期,因为相互之间还有些许连接;图4中染色体个数约为10,少于40个。




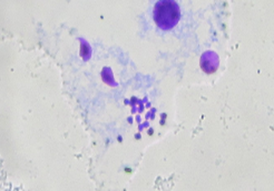
图5小鼠精细胞(方框)及中期染色体(圈) 图6:中期染色体(圈)及其他细胞(10*40)
【分析】这两幅图的中期染色体都较为清晰,图5为14条,图6为20条;图6颜色较紫;
2、实验结果分析
A、由图6的标注可以看出,精细胞是一端较为尖细的细胞,较尖的一端将来会长出精子尾结构;中期染色体彼此并不连接,而在视野中还有处于其他状态下的细胞;如果有完整的细胞,详细辨认可能看到的细胞种类有:精原细胞——是精子发生的干细胞。胞体圆形,核居中央,染色质呈疏松网状结构;初级精母细胞——进行第一次减数分裂的细胞,由精原细胞分裂产生,胞体较大,浅染,偶见染色质丝;次级精母细胞——由初级精母细胞分裂而来,胞体较初级精母细胞小,染色质呈细网状,着色较浅;精细胞——体积较次级精母细胞更小,着色深,多呈球形或精子头的雏形;精子——形态不同于精细胞,主体梭形,带一小段尾巴。
B、图2及图3中的染色体个数为64条,大于小鼠体细胞中的染色体个数;图4为10条,图5为14条,图6为20条;
C、根据染色体条数,可以基本判断,图2及图3处于有丝分裂的中期,或者是减数第一次分裂的中期;图6可能是处于减数第二次分裂的中期;
D、以上确定的依据是:小鼠正常的体细胞的染色体数为40条,若超过40条,分析造成这种结果的原因为几个细胞的染色体聚集到一起;在减数第二次分裂中期,由于减数第一次分裂使得染色体的数目减半成为20条,因此若没有染色体的丢失,图6所示的染色体为减数第二次分裂中期的染色体;而由于实验操作中,低渗处理,离心以及制片时滴细胞悬液的高度问题,一组染色体或许会被摔碎,变得不完整,出现图4,图5的情况;或许几个细胞的染色体混在一起,出现图2图3的情况;
E、由于制片时细胞悬液吹打不均匀,细胞悬液滴下的高度不够,在桌边震荡不够等等的操作原因,都可能够造成细胞的分散不够彻底,大团的细胞集中在一起,难以观察的现象,就像图4所示,在一定程度上影响观察;
F、若实验过程中低渗的时间过长,细胞将会破裂严重,在离心之前,细胞就已经破裂,更难以观察中期的染色体,估计图2中背景模糊混乱,与低渗过度,细胞不正常破裂有关;
【实验反思与总结】
1、实验注意事项
① 注射秋水仙素:应选择腹腔注射,如果观察到小鼠被注射后皮肤隆起,则表明注射到皮下,应重新注射;如果在拔出针时观察到有血珠溢出,则是因为注射时损伤了部分器官或血管,秋水仙素会随血液传递到全身,其传递速度会加快,其死亡时间可能小于15-16h,应随时观察小鼠,以防其突然死亡。即使注射到
小鼠的腹腔中,也很有可能注射到起脂肪组织中,使小鼠的死亡时间变慢。因此应根据具体实验,适当调整注射时间和等待时间。秋水仙素的注射浓度过高或者处理时间过长会造成染色体的过度凝集,不利于形态的观察;
② 处死小鼠:最好选择在小鼠将要死亡时处死小鼠,因为此时,秋水仙素的作用发挥到最大,分裂期中期的细胞最多;如果等小鼠死亡后处死小鼠,小鼠尸体可能已僵硬,难以取到精巢。
③ 剪碎组织:得到睾丸时,要尽量剥干净被膜和结缔组织,可以取少于1mL0.3% KCl溶液,没过杯底即可,如果液体太多则很难剪到组织,比较浪费时间。
④ 铜网过滤:应把组织尽量剪碎后再用铜网过滤;如果烧杯中仍有较大颗粒,则小心不要把那些颗粒倒出,在烧杯中再加入少量0.3% KCl溶液,继续剪;如果铜网上还有很多比较大的颗粒,先用少量0.3% KCl溶液将其冲入烧杯,再进行剪碎。
⑤ 实验步骤中的(4)所提到的30mins包括包括(5)中的离心配平时间,因此,
真正的静置时间应少于30mins,细胞与0.3% KCl接触的总时间应为45mins左右。另外,如果在最后的镜检时,观察到细胞呈圆形,比较膨胀,即证明低渗这一步骤的成功。反之,如果低渗不成功,则细胞会比较瘪,且这样的细胞很难摔碎。 低渗处理的目的是使细胞的体积膨大,染色体松散。处理时间过长则会使细胞破裂,染色体丢失,时间过短则染色体不能够良好分散;
⑥ 离心:离心的转速是800-1000r/min,这样低的转速是为了防止细胞破裂。
⑦ 再次离心和固定:这样做是因为第一次离心之前细胞经过了低渗处理,虽然抽去了上清液,但是不可能抽干净,因此,再次离心保证细胞离开低渗环境。
⑧ 吹散细胞:应该轻轻地吹细胞,如果太用力会导致细胞破裂。
⑨ 摔碎细胞:摔碎细胞时应注意滴管与载玻片的距离,如果距离太小,则细胞很难摔碎,不会观察到染色体;但如果距离太大,会使细胞完全的碎开,染色体布满视野,虽然观察到染色体,但不能明确分出哪些是同一个细胞的,会给计数造成一定的难度。
⑩ 实际操作发现,细胞很难摔碎,这可以归结为多方面原因。首先,如果在剪碎组织时没有剪好,则会导致细胞匀浆中含有大量细胞团,由于这些细胞聚集在一起,就有一定的组织强度,十几厘米的高度很难摔碎,因此,这一步骤十分关键。然后,如果低渗不完全,细胞可能膨胀程度不够,很难摔碎。再次,如果混匀细胞时没有完全混匀,则会导致细胞聚集在一起,敲细胞时很难敲碎,同样观察不到理想的效果;如果吹得太用力,则会导致细胞被吹碎,视野中有大量细胞碎片,影响观察。当然,如果摔细胞时高度不够,细胞不可能被摔碎。
? 染色:染色之后应冲去染液并晾干,注意使用小水流并从背面冲洗,这样可以避免冲去细胞。
2、补充:染色体疾病
染色体数目或结构异常引起的疾病称为染色体病(chromosomal disorder)。这类疾病的实质是染色体上的基因或基因群的增减或变位影响了众多基因的表达和作用,严重地破坏了基因的平衡状态,因而妨碍了人体相关器官的分化发育,造成机体形态和功能的异常。严重者在胚胎早期夭折并引起自发流产,故染色体异常易见于自发流产胎儿。少数即使能存活到出生,也往往表现有生长、智力或性发育异常和先天性多发畸形。因此,染色体病对人类危害甚大,且又无治疗良策,目前主要通过遗传咨询和产前诊断予以预防。染色体病表型的轻重程度主要取决于染色体上所累及基因的数量和功能。
染色体病按染色体种类和表型可分为三种:常染色体病、性染色体病和染色体异常的携带者。染色体病在临床上和遗传上一般有如下特点:①染色体病患者均有先天性多发畸形(包括特殊面容)、生长、智力或性发育落后、特殊肤纹;②绝大多数染色体病患者呈散发性,即双亲染色体正常,畸变染色体来自双亲生殖细胞或受精卵早期卵裂新发生的染色体畸变,这类患者往往无家族史;③少数染色体结构畸变的患者是由表型正常的双亲遗传而得,其双亲之一为平衡的染色体结构重排携带者,可将畸变的染色体遗传给子代,引起子代的染色体不平衡而致病,这类患者常伴有家族史。
3、实验小结
此次实验一共耗时两周,第一周制片,第二周染色及观察;第一周实验过程中由于低渗的时间有些长,因此细胞破裂较多,致使制成的细胞悬液制成装片后背景杂乱,染色体不完整;在染色时,中间部分有些地方有气泡,因此染色不够充分,或许还有细胞的破裂等等原因,有些地方的染色稍微显紫红色;在拍照时,焦距并没有调好,因此照片不是很清晰。但是找到了中期的染色体,并且看到了精细胞及其他细胞,实验基本成功;
在此次实验中我学到做事要耐心细致,因为拿玻片时不小心,在此次实验中将玻片打碎了一点,影响观察;在找中期染色体时,开始时出现了不耐心的现象,但在后期,耐下心来还是找到了中期的染色体,相信自己,坚持细致,才能够取得实验的成功。
第二篇:细胞生物学实验报告1- 动物细胞融合
【实验目的】
1.了解动物细胞融合的常用方法
2.学习化学融合和电融合的基本操作
3.观察动物细胞融合过程中的行为和变化
【实验原理】
1.病毒诱导融合
仙台病毒、牛痘病毒、新城鸡瘟病毒和疱疹病毒等可以介导细胞的融合。这类病毒的被膜中含有融合蛋白,可以介导病毒同宿主细胞融合,也可以介导细胞与细胞的融合。用紫外线灭活后,这些病毒即可诱导细胞发生融合。
2.化学诱导融合
很多化学试剂都能诱导细胞融合,如聚乙二醇(PEG),二甲基亚砜、山梨醇、甘油、溶血性卵磷脂,磷脂酰丝氨酸等。这些物质能够改变细胞膜脂质分子的排列,在去除这些物质之后,细胞膜趋向于恢复原有的有序结构。在恢复的过程中相接处的细胞由于接口处脂质双分子层的相互亲和与表面张力,细胞膜融合,胞质流通,发生融合。化学诱导方法,操作方便,诱导融合的概率比较高,效果稳定,适用于动、植物细胞,但对细胞具有一定的毒性。PEG是被广泛使用的化学融合剂。
3.电击诱导融合
包括电诱导、激光诱导等。其中,电诱导是先使细胞在电场中极化成为偶极子,沿电力线排布成串,再利用高强度、短时程的电脉冲击破细胞膜,细胞膜的脂质分子发生重排,由于表面张力作用,两细胞发生融合。电诱导方法具有融合过程易控制,融合概率高,作用机制明确,可重复性高等优点。
【实验材料】
1.材料 鸡血红细胞
2.试剂 50%PEG、0.85%氯化钠溶液、GKN缓冲溶液
3.器材 倒置显微镜、离心机、量筒、注射器、载玻片、盖玻片、离心管等
【实验步骤】
1.取离心管,加入冷藏的鸡血1ml,加入4ml 0.85%氯化钠溶液,摇匀
2.将离心管放入离心机中,以1000-1200r/min的速度离心5分钟左右。取出离心管,弃掉上清液,再加入4ml氯化钠溶液,重复以上操作。
3.取离心管,弃掉上清液后,按照红细胞沉积的体积,加入1-2mlGKN溶液,摇匀。
4.取以上溶液1ml,加入3mlGKN溶液,再次摇匀
5.取以上溶液1ml,加入0.5mlPEG,先静置1-2min,再将溶液摇匀后,静置1-2min。
6.用滴管取溶液制成涂片,置于显微镜下观察。
【实验结果】
最初,在显微镜下观察到的细胞如下二图所示。
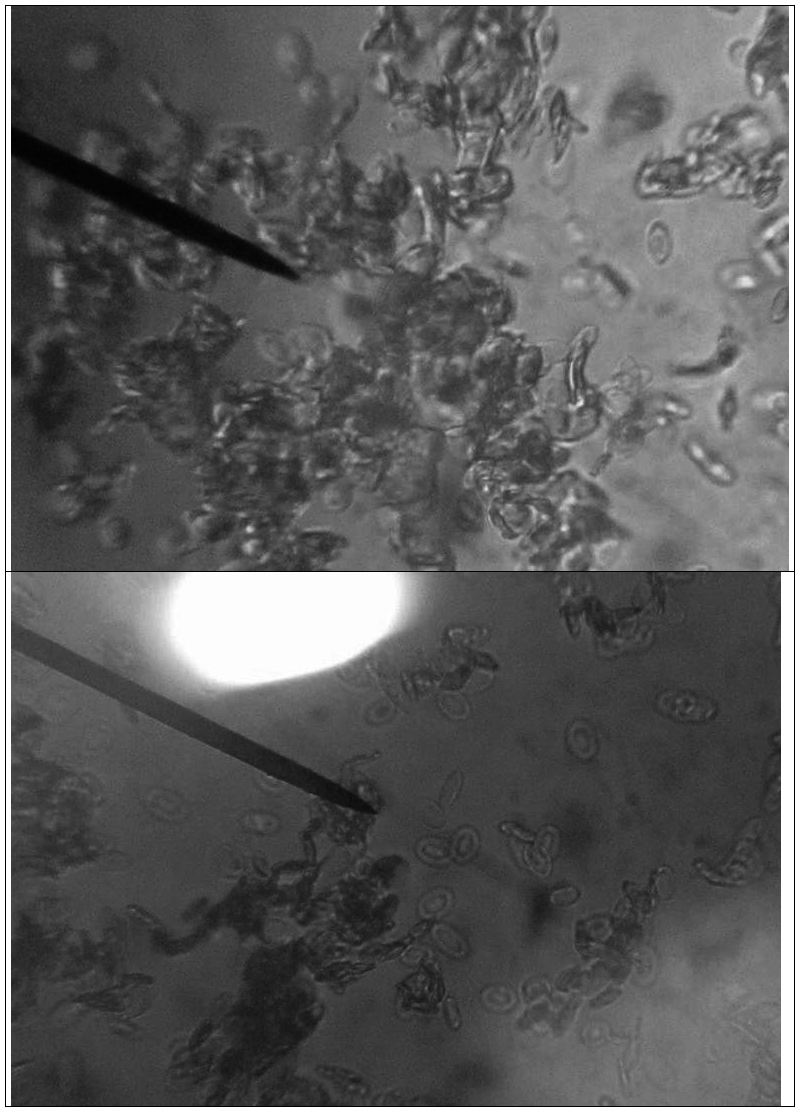
其中,第二图中指针所指的两细胞紧贴在一起,有相互融合的趋势。由于在第二步稀释的过程中未充分振荡,使得许多细胞都聚集在一起成堆,影响了细胞的观察。因此,我又重新充分稀释并振荡了细胞悬液,并重新制备了涂片,所得的新影响如下图:

在上图中,可以明显看出箭头所指的两细胞中间的细胞膜已经消失。有着明显的融合的现象了。
【实验结果分析与注意事项】
本次试验最终观察到了明显的细胞融合,可以说较为成功。其中重新稀释振荡细胞悬液并重置涂片,使得能够看到明显的细胞融合现象。而其他实验组也出现了类似情况,都是细胞悬液稀释程度不够,制成的图片在显微镜下呈现大块的细胞堆积,难以观察到明显而清晰的单个细胞,更不用说两个细胞的融合了。因此在本次试验中,第一个注意事项便是充分地稀释和振荡摇匀细胞悬液,达到试验的细胞浓度要求(17000个/毫升左右)。
许多实验组由于在制备的第五步(滴入PEG)过程中,滴入PEG后立刻摇匀了细胞悬液,并没有让PEG沉降到细胞悬液底部后,在保持高浓度的情况下与接触面的细胞充分反应,使得所观察到了细胞融合率不高,观察效果不明显。因此,让PEG与细胞悬液分层静置足够的时间也是十分重要的。
另外,包括我组在内的许多实验组在观察细胞是发现目标细胞总是在不规则移动,十分影响细胞的观察。根据指导老师的说法,这是由于涂片的悬液量太多导致细胞在悬液内流动所致。因此,在制片时,适当的悬液量是需要注意的。不能过多,当然也不能过少。
附:
细胞融合技术的应用
1.单克隆抗体
动物脾脏有上百万种不同的B淋巴细胞系,具有不同基因不同的B淋巴细胞合成不同的抗体。当机体受抗原刺激时,抗原分子上的许多决定簇分别激活各个具有不同基因的B细胞。被激活的B细胞分裂增殖形成效应B细胞(浆细胞)和记忆B细胞,大量的浆细胞克隆合成和分泌大量的抗体分子分布到血液、体液中。如果能选出一个制造一种专一抗体的浆细胞进行培养,就可得到由单细胞经分裂增殖而形成细胞群,即单克隆。单克隆细胞将合成针对一种抗原决定簇的抗体,称为单克隆抗体。
1975年分子生物学家G.J.F.克勒和C.米尔斯坦在细胞杂交技术的基础上,创建立杂交瘤技术,他们把可在体外培养和大量增殖的小鼠骨髓瘤细胞与经抗原免疫后的纯系小鼠脾细胞融合,成为杂交细胞系,既具有瘤细胞易于在体外无限增殖的特性,又具有抗体形成细胞的合成和分泌特异性抗体的特点。将这种杂交瘤作单个细胞培养,可形成单细胞系,即单克隆。利用培养或小鼠腹腔接种的方法,便能得到大量的、高浓度的、非常均一的抗体,其结构、氨基酸顺序、特异性等都是一致的,而且在培养过程中,只要没有变异,不同时间所分泌的抗体都能保持同样的结构与机能。这种单克隆抗体是用其他方法所不能得到的。这项新技术从根本上解决了在抗体制备中长期存在的特异性和可重复性问题,可用于探讨①蛋白质的精细结构;②淋巴细胞亚群的表面新抗原;③组织相容性抗原;④激素和药物的放射免疫(或酶免疫)分析;⑤肿瘤的定位和分类;⑥纯化微生物和寄生虫抗原;⑦免疫治疗和与药物结合的免疫-化学疗法 (“导弹”疗法,利用单克隆抗体与靶细胞特异性结合,将药物带至病灶部位。因此,单克隆抗体可直接用于人类疾病的诊断、预防、治疗以及免疫机制的研究,为人类恶性肿瘤的免疫诊断与免疫治疗开辟了广阔前景。
2.植物细胞融合育种
植物细胞融合可分为体细胞杂交和配子-体细胞杂交,前者是指不经过有性生殖,而直接由体细胞原生质体融合产生杂种细胞,形成愈伤组织,并再生出植株的过程,后者是指性细胞原生质体和二倍体原生质体融合产生三倍体杂种细胞,形成愈伤组织,并再生出植株的过程。植物细胞融合是植物细胞工程的一个重要分支,是一种突破物种生殖隔离、创造远缘杂交的新途径。
自1960年Cocking用酶解法分离出番茄根原生质体后,Nataga和Takebe1970年首次利用烟草叶分离出原生质体,经培养获得再生植株;1975年以色列的Vardi等首次从木本植物shamonti甜橙珠心组织诱导胚愈伤组织,并从愈伤组织分离原生质体,经培养通过胚状体再生出植株;在禾本科植物中,除灾珍珠谷、紫狼尾草用悬浮细胞为材料,较早获得原生质体再生植株外,直到1985年Fujimura等率先在水稻原生质体培养中获得了再生植株,才出现了重大突破。现在已从许多种内、种间、属间甚至亚科间的体细胞杂交获得杂交细胞系或杂种植物。随着多种细胞原生质体的成功培养和融合技术的不断改进,植物细胞融合获得了巨大成功。目前已经成功得到的融合植株有甘蓝-青菜、大豆-马塘草、矮牵牛-龙面花、大麦-花生、大麦-大豆、小麦-矮牵牛、油菜-大豆、玉米-大豆、大豆-野豌豆、大麦-蚕豆、大豆-香草木樨、大豆-烟草和大豆-秋水仙等。
3.微生物原生质体融合构成新菌株
微生物原生质体的获得、纯化、培养等类似于植物原生质体。微生物原生质体融合是一种方法简便用途较广的技术,在育种中将会有更多的实际应用。
-
细胞生物学综合性实验报告
本科学生实验报告学号姓名学院生命科学学院专业班级11级生物科学C班实验课程名称细胞生物学实验指导教师及职称吴暇玉开课时间20xx至…
-
细胞生物学实验报告
染色体标本的制备及观察泮力菁20xx0014009120xx级生物基地同组者商倩倩实验目的1掌握染色体标本制作的方法2认识不同生物…
-
细胞生物学实验报告
细胞生物学实验报告细胞生物学实验报告叶绿体的分离纯化与荧光观察实验目的通过植物细胞叶绿体的分离与纯化了解细胞器分离与纯化的原理和方…
-
细胞生物学综合实验报告
本科学生实验报告姓名学院生命科学学院专业班级10B班实验课程名称人类淋巴细胞株的培养指导教师及职称许琳峰开课时间20xx至20xx…
-
细胞生物学实验报告
细胞生物学实验报告细胞生物学实验报告小鼠脾细胞原代培养姓名杜旭学号20xx00230155系年级20xx级临七一班同组者赵伟日期2…
-
动物细胞培养助教总结报告
动物细胞培养助教总结报告李骥3009213008一实验目的1了解动物细胞培养的概念与分类2熟悉无菌操作的步骤以及技巧二实验原理1动…
-
细胞生物学综合性实验报告
本科学生实验报告学号姓名学院生命科学学院专业班级11级生物科学C班实验课程名称细胞生物学实验指导教师及职称吴暇玉开课时间20xx至…
-
原代细胞培养实验报告
实验细胞培养1实验目的初步掌握哺乳动物细胞的原代培养与传代培养的基本操作过程为生物工程在医学上的应用打下基础2实验原理从生物体中取…
-
细胞生物学实验报告
细胞生物学实验报告细胞生物学实验报告小鼠脾细胞原代培养姓名杜旭学号20xx00230155系年级20xx级临七一班同组者赵伟日期2…
-
细胞生物学实验报告5
湖南科技学院实验报告学生姓名李诗学号20xx07004215专业生物技术年级班级1102课程名称细胞生物学实验实验指导老师蒋琼凤沈…
-
细胞工程实验报告
细胞工程实验报告专业生物技术班级0801姓名励丹学号30804305一实验目的1了解动物细胞培养的相关实验器材以及实验前器材的清洗…